

Viruses and the autophagy pathway
- PMID: 25858140
- PMCID: PMC5917100
- DOI: 10.1016/j.virol.2015.03.042
Viruses and the autophagy pathway
Abstract
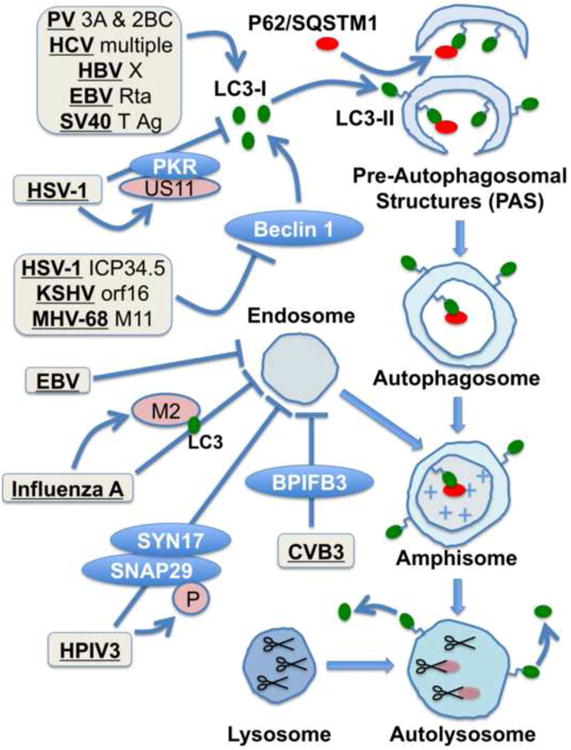
Studies of the cellular autophagy pathway have exploded over the past twenty years. Now appreciated as a constitutive degradative mechanism that promotes cellular homeostasis, autophagy is also required for a variety of developmental processes, cellular stress responses, and immune pathways. Autophagy certainly acts as both an anti-viral and pro-viral pathway, and the roles of autophagy depend on the virus, the cell type, and the cellular environment. The goal of this review is to summarize, in brief, what we know so far about the relationship between autophagy and viruses, particularly for those who are not familiar with the field. With a massive amount of relevant published data, it is simply not possible to be comprehensive, or to provide a complete "parade of viruses", and apologies are offered to researchers whose work is not described herein. Rather, this review is organized around general themes regarding the relationship between autophagy and animal viruses.
Keywords: AWOL; Amphisome; Autophagy; Beclin 1; Endosome; IFN; SNARE; Virus.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
