

Substrate degradation by the proteasome: a single-molecule kinetic analysis
- PMID: 25859050
- PMCID: PMC4450770
- DOI: 10.1126/science.1250834
Substrate degradation by the proteasome: a single-molecule kinetic analysis
Abstract
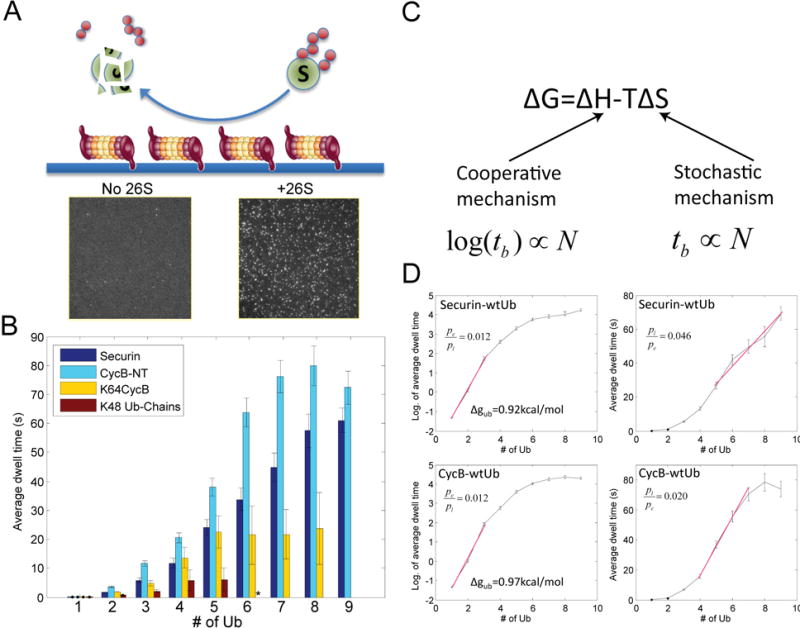
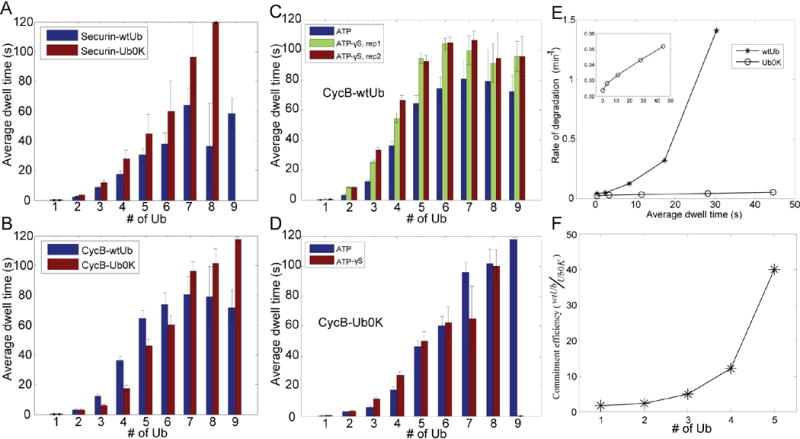
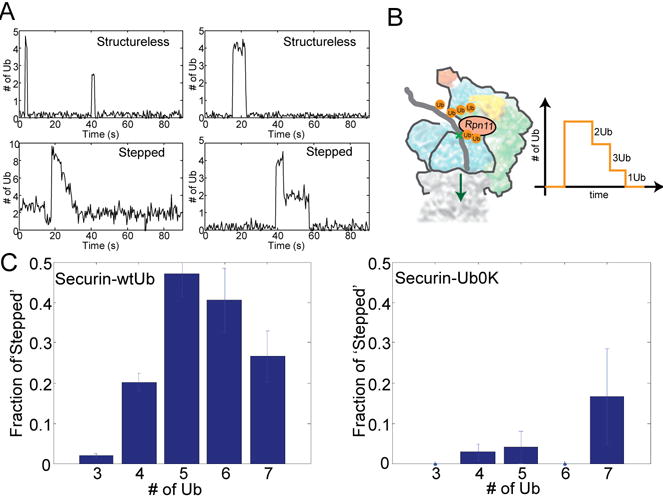
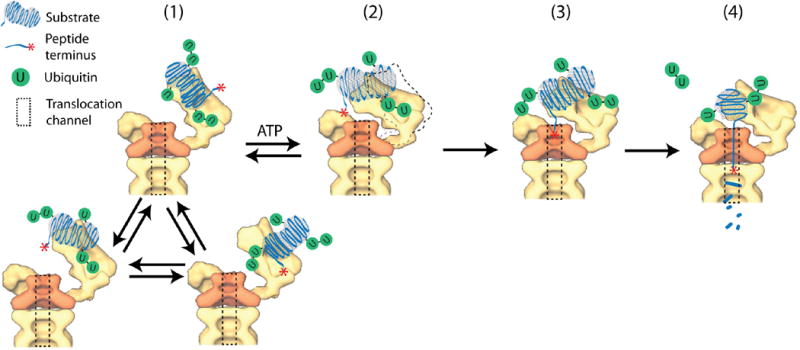
To address how the configuration of conjugated ubiquitins determines the recognition of substrates by the proteasome, we analyzed the degradation kinetics of substrates with chemically defined ubiquitin configurations. Contrary to the view that a tetraubiquitin chain is the minimal signal for efficient degradation, we find that distributing the ubiquitins as diubiquitin chains provides a more efficient signal. To understand how the proteasome actually discriminates among ubiquitin configurations, we developed single-molecule assays that distinguished intermediate steps of degradation kinetically. The level of ubiquitin on a substrate drives proteasome-substrate interaction, whereas the chain structure of ubiquitin affects translocation into the axial channel on the proteasome. Together these two features largely determine the susceptibility of substrates for proteasomal degradation.
Copyright © 2015, American Association for the Advancement of Science.
Figures

Comment in
-
Biochemistry. Details of destruction, one molecule at a time.Science. 2015 Apr 10;348(6231):183-4. doi: 10.1126/science.aab0931. Science. 2015. PMID: 25859031 No abstract available.
References
-
- Pickart CM, Eddins MJ. Ubiquitin: structures, functions, mechanisms. Biochim Biophys Acta. 2004 Nov 29;1695:55. - PubMed
-
- Wilkinson KD. Regulation of ubiquitin-dependent processes by deubiquitinating enzymes. FASEB J. 1997 Dec;11:1245. - PubMed
-
- Kirkpatrick DS, et al. Quantitative analysis of in vitro ubiquitinated cyclin B1 reveals complex chain topology. Nat Cell Biol. 2006 Jul;8:700. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
