

Oxidative stress and starvation in Dinoroseobacter shibae: the role of extrachromosomal elements
- PMID: 25859246
- PMCID: PMC4373377
- DOI: 10.3389/fmicb.2015.00233
Oxidative stress and starvation in Dinoroseobacter shibae: the role of extrachromosomal elements
Abstract
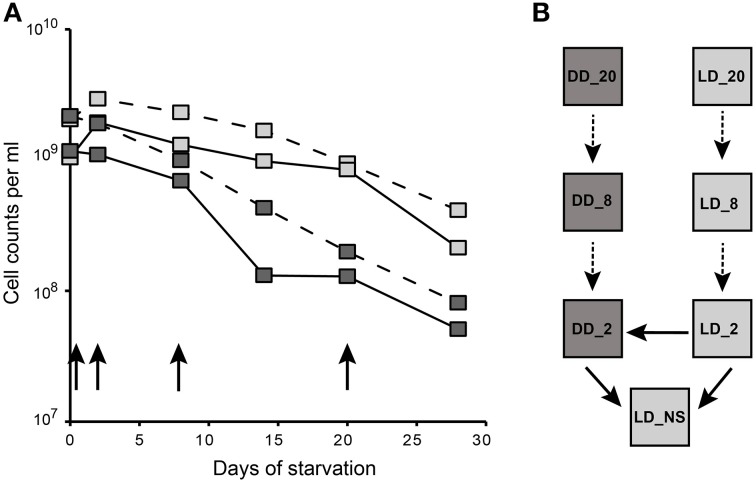
Aerobic anoxygenic phototrophic bacteria (AAP) are abundant in the photic zone of the marine environment. Dinoroseobacter shibae, a representative of the Roseobacter group, converts light into additional energy that enhances its survival especially under starvation. However, light exposure results in the production of cytotoxic reactive oxygen species in AAPs. Here we investigated the response of D. shibae to starvation and oxidative stress, focusing on the role of extrachromosomal elements (ECRs). D. shibae possessing five ECRs (three plasmids and two chromids) was starved for 4 weeks either in the dark or under light/dark cycles and the survival was monitored. Transcriptomics showed that on the chromosome genes with a role in oxidative stress response and photosynthesis were differentially expressed during the light period. Most extrachromosomal genes in contrast showed a general loss of transcriptional activity, especially in dark-starved cells. The observed decrease of gene expression was not due to plasmid loss, as all five ECRs were maintained in the cells. Interestingly, the genes on the 72-kb chromid were the least downregulated, and one region with genes of the oxygen stress response and a light-dependent protochlorophyllide reductase of cyanobacterial origin was strongly activated under the light/dark cycle. A Δ72-kb curing mutant lost the ability to survive under starvation in a light/dark cycle demonstrating the essential role of this chromid for adaptation to starvation and oxidative stress. Our data moreover suggest that the other four ECRs of D. shibae have no vital function under the investigated conditions and therefore were transcriptionally silenced.
Keywords: aerobic anoxygenic photosynthesis; chlorophyll a biosynthesis; light-stress adaptation; plasmid-maintenance; reactive oxygen species; starvation.
Figures






Similar articles
-
Transcriptional response of the photoheterotrophic marine bacterium Dinoroseobacter shibae to changing light regimes.ISME J. 2011 Dec;5(12):1957-68. doi: 10.1038/ismej.2011.68. Epub 2011 Jun 9. ISME J. 2011. PMID: 21654848 Free PMC article.
-
Light enhances survival of Dinoroseobacter shibae during long-term starvation.PLoS One. 2013 Dec 30;8(12):e83960. doi: 10.1371/journal.pone.0083960. eCollection 2013. PLoS One. 2013. PMID: 24386315 Free PMC article.
-
Characteristics and Evolutionary Analysis of Photosynthetic Gene Clusters on Extrachromosomal Replicons: from Streamlined Plasmids to Chromids.mSystems. 2019 Sep 10;4(5):e00358-19. doi: 10.1128/mSystems.00358-19. mSystems. 2019. PMID: 31506262 Free PMC article.
-
Extrachromosomal, extraordinary and essential--the plasmids of the Roseobacter clade.Appl Microbiol Biotechnol. 2013 Apr;97(7):2805-15. doi: 10.1007/s00253-013-4746-8. Epub 2013 Feb 24. Appl Microbiol Biotechnol. 2013. PMID: 23435940 Review.
-
The trouble with oxygen: The ecophysiology of extant phototrophs and implications for the evolution of oxygenic photosynthesis.Free Radic Biol Med. 2019 Aug 20;140:233-249. doi: 10.1016/j.freeradbiomed.2019.05.003. Epub 2019 May 9. Free Radic Biol Med. 2019. PMID: 31078729 Review.
Cited by
-
Adaptation of Dinoroseobacter shibae to oxidative stress and the specific role of RirA.PLoS One. 2021 Mar 29;16(3):e0248865. doi: 10.1371/journal.pone.0248865. eCollection 2021. PLoS One. 2021. PMID: 33780465 Free PMC article.
-
Plasmid-Mediated Stabilization of Prophages.mSphere. 2022 Apr 27;7(2):e0093021. doi: 10.1128/msphere.00930-21. Epub 2022 Mar 21. mSphere. 2022. PMID: 35311569 Free PMC article.
-
The Sixth Element: a 102-kb RepABC Plasmid of Xenologous Origin Modulates Chromosomal Gene Expression in Dinoroseobacter shibae.mSystems. 2022 Aug 30;7(4):e0026422. doi: 10.1128/msystems.00264-22. Epub 2022 Aug 3. mSystems. 2022. PMID: 35920548 Free PMC article.
-
A novel roseosiphovirus infecting dinoroseobacter shibae DFL12T represents a new genus.BMC Genomics. 2025 Feb 8;26(1):121. doi: 10.1186/s12864-025-11274-w. BMC Genomics. 2025. PMID: 39923004 Free PMC article.
-
Transcriptome and Deletion Mutant Analyses Revealed that an RpoH Family Sigma Factor Is Essential for Photosystem Production in Roseateles depolymerans under Carbon Starvation.Microbes Environ. 2023;38(1):ME22072. doi: 10.1264/jsme2.ME22072. Microbes Environ. 2023. PMID: 36878600 Free PMC article.
References
-
- Benjamini Y., Hochberg Y. (1995). Controlling the false discovery rate: a practical and powerful approach to multiple testing. J. R. Stat. Soc. 57, 289–300.
-
- Berger M., Brock N. L., Liesegang H., Dogs M., Preuth I., Simon M., et al. (2012). Genetic analysis of the upper phenylacetate catabolic pathway in the production of tropodithietic acid by Phaeobacter gallaeciensis. Appl. Microbiol. Biotechnol. 78, 3539–3551 10.1128/AEM.07657-11 - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous