

Expression and function of S100A8/A9 (calprotectin) in human typhoid fever and the murine Salmonella model
- PMID: 25860480
- PMCID: PMC4393299
- DOI: 10.1371/journal.pntd.0003663
Expression and function of S100A8/A9 (calprotectin) in human typhoid fever and the murine Salmonella model
Abstract
Background: Typhoid fever, caused by the Gram-negative bacterium Salmonella enterica serovar Typhi, is a major cause of community-acquired bacteremia and death worldwide. S100A8 (MRP8) and S100A9 (MRP14) form bioactive antimicrobial heterodimers (calprotectin) that can activate Toll-like receptor 4, promoting lethal, endotoxin-induced shock and multi-organ failure. We aimed to characterize the expression and function of S100A8/A9 in patients with typhoid fever and in a murine invasive Salmonella model.
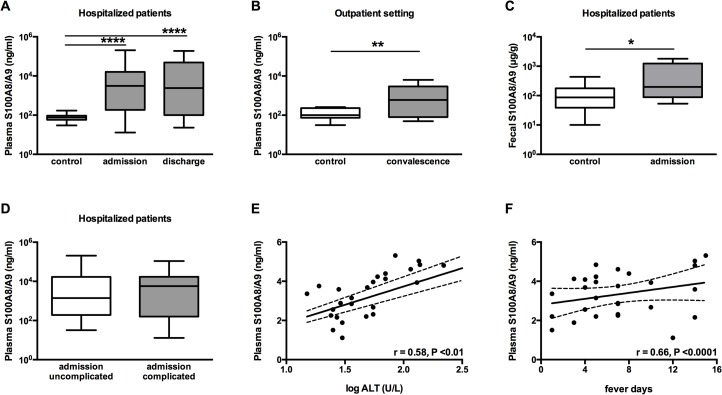
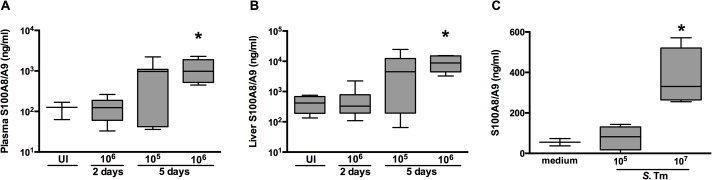
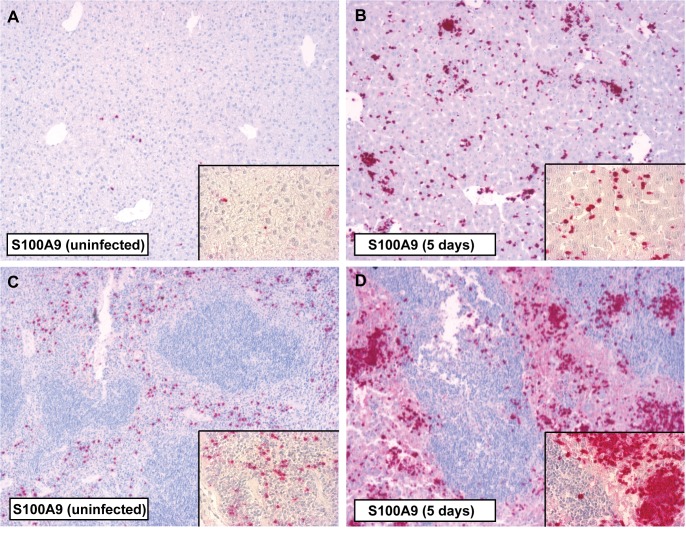
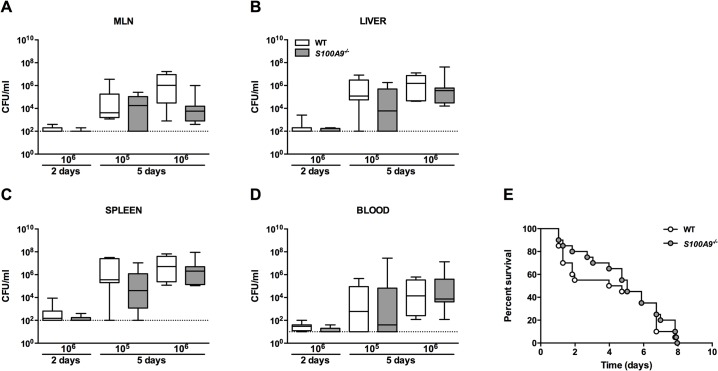
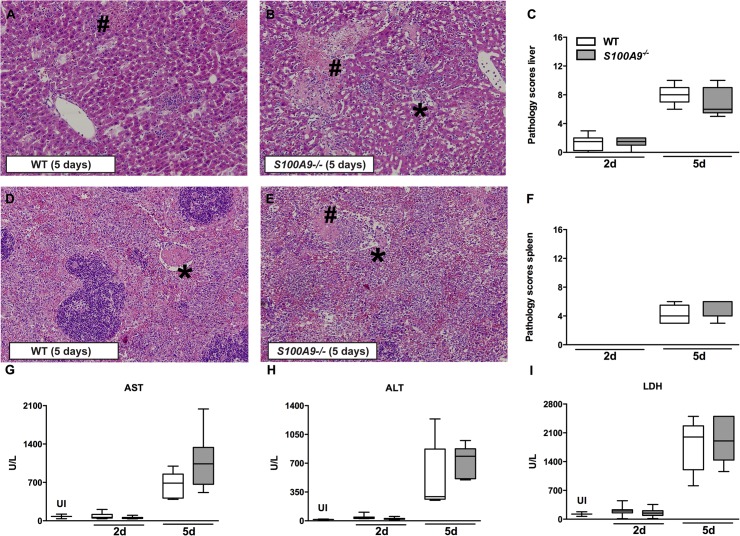
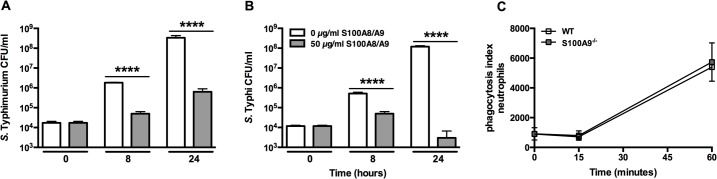
Methods and principal findings: S100A8/A9 protein levels were determined in acute phase plasma or feces from 28 Bangladeshi patients, and convalescent phase plasma from 60 Indonesian patients with blood culture or PCR-confirmed typhoid fever, and compared to 98 healthy control subjects. To functionally characterize the role of S100A8/A9, we challenged wildtype (WT) and S100A9-/- mice with S. Typhimurium and determined bacterial loads and inflammation 2- and 5- days post infection. We further assessed the antimicrobial function of recombinant S100A8/A9 on S. Typhimurium and S. Typhi replication in vitro. Typhoid fever patients demonstrated a marked increase of S100A8/A9 in acute phase plasma and feces and this increases correlated with duration of fever prior to admission. S100A8/A9 directly inhibited the growth of S. Typhimurium and S. Typhi in vitro in a dose and time dependent fashion. WT mice inoculated with S. Typhimurium showed increased levels of S100A8/A9 in both the liver and the systemic compartment but S100A9-/- mice were indistinguishable from WT mice with respect to bacterial growth, survival, and inflammatory responses, as determined by cytokine release, histopathology and organ injury.
Conclusion: S100A8/A9 is markedly elevated in human typhoid, correlates with duration of fever prior to admission and directly inhibits the growth of S. Typhimurium and S. Typhi in vitro. Despite elevated levels in the murine invasive Salmonella model, S100A8/A9 does not contribute to an effective host response against S. Typhimurium in mice.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Parry CM, Hien TT, Dougan G, White NJ, Farrar JJ (2002) Typhoid fever. N Engl J Med 347: 1770–1782. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
