

Induction of microglia activation after infection with the non-neurotropic A/CA/04/2009 H1N1 influenza virus
- PMID: 25861024
- PMCID: PMC4393251
- DOI: 10.1371/journal.pone.0124047
Induction of microglia activation after infection with the non-neurotropic A/CA/04/2009 H1N1 influenza virus
Abstract
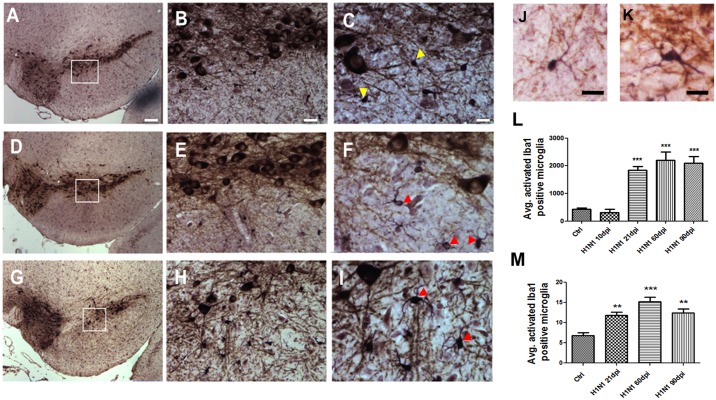
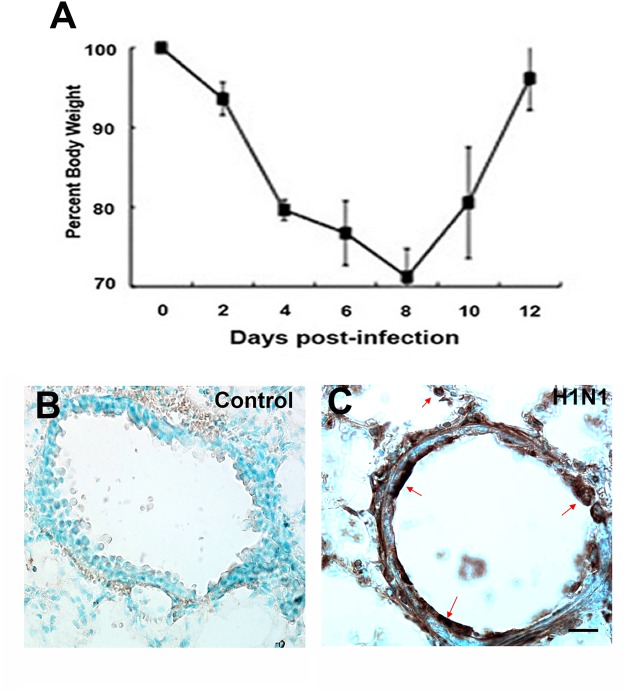
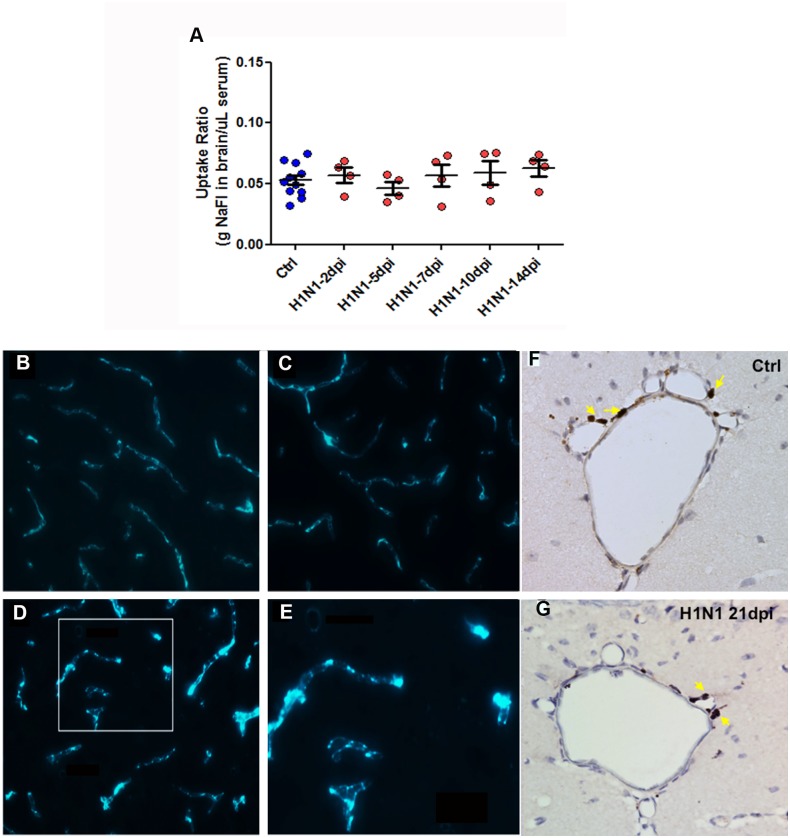
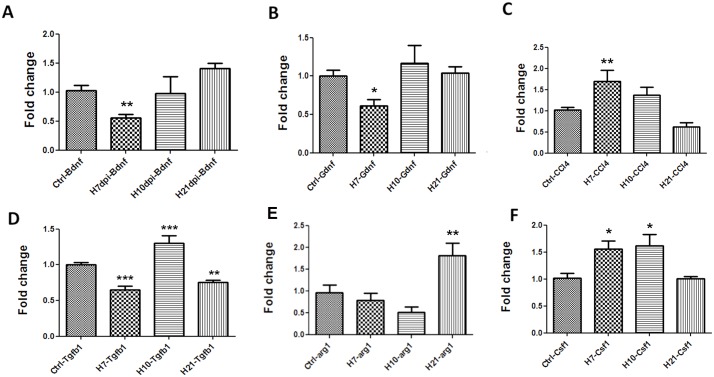
Although influenza is primarily a respiratory disease, it has been shown, in some cases, to induce encephalitis, including people acutely infected with the pandemic A/California/04/2009 (CA/09) H1N1 virus. Based on previous studies showing that the highly pathogenic avian influenza (HPAI) A/Vietnam/1203/2004 H5N1 virus was neurotropic, induced CNS inflammation and a transient parkinsonism, we examined the neurotropic and inflammatory potential of the CA/09 H1N1 virus in mice. Following intranasal inoculation, we found no evidence for CA/09 H1N1 virus neurotropism in the enteric, peripheral or central nervous systems. We did, however, observe a robust increase in microglial activity in the brain characterized by an increase in the number of activated Iba-1-positive microglia in the substantia nigra (SN) and the hippocampus, despite the absence of virus in the brain. qPCR analysis in SN tissue showed that the induction of microgliosis was preceded by reduced gene expression of the neurotrophic factors bdnf, and gdnf and increases in the immune modulatory chemokine chemokine (C-C motif) ligand 4 (ccl4). We also noted changes in the expression of transforming growth factor-1 (tgfβ1) in the SN starting at 7 days post-infection (dpi) that was sustained through 21 dpi, coupled with increases in arginase-1 (arg1) and csf1, M2 markers for microglia. Given that neuroinflammation contributes to generation and progression of a number of neurodegenerative disorders, these findings have significant implications as they highlight the possibility that influenza and perhaps other non-neurotropic viruses can initiate inflammatory signals via microglia activation in the brain and contribute to, but not necessarily be the primary cause of, neurodegenerative disorders.
Conflict of interest statement
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
