

Loss of a Clueless-dGRASP complex results in ER stress and blocks Integrin exit from the perinuclear endoplasmic reticulum in Drosophila larval muscle
- PMID: 25862246
- PMCID: PMC4434815
- DOI: 10.1242/bio.201511551
Loss of a Clueless-dGRASP complex results in ER stress and blocks Integrin exit from the perinuclear endoplasmic reticulum in Drosophila larval muscle
Abstract
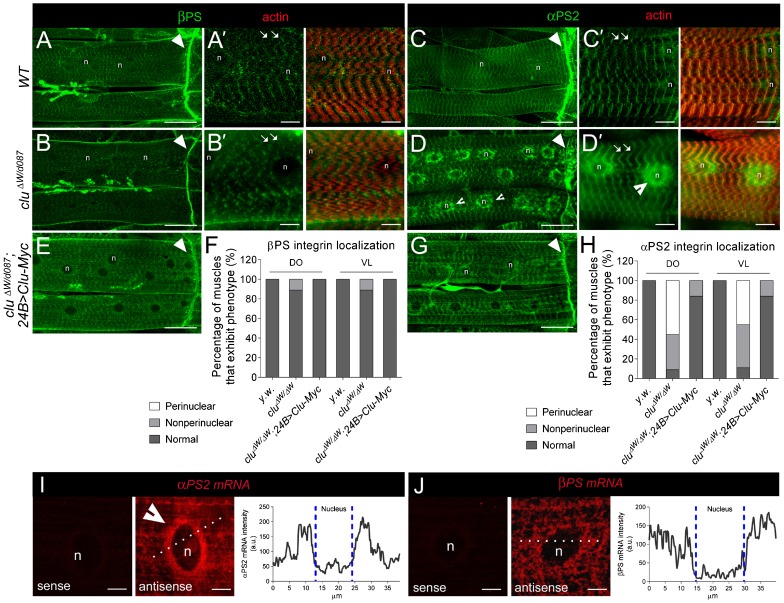
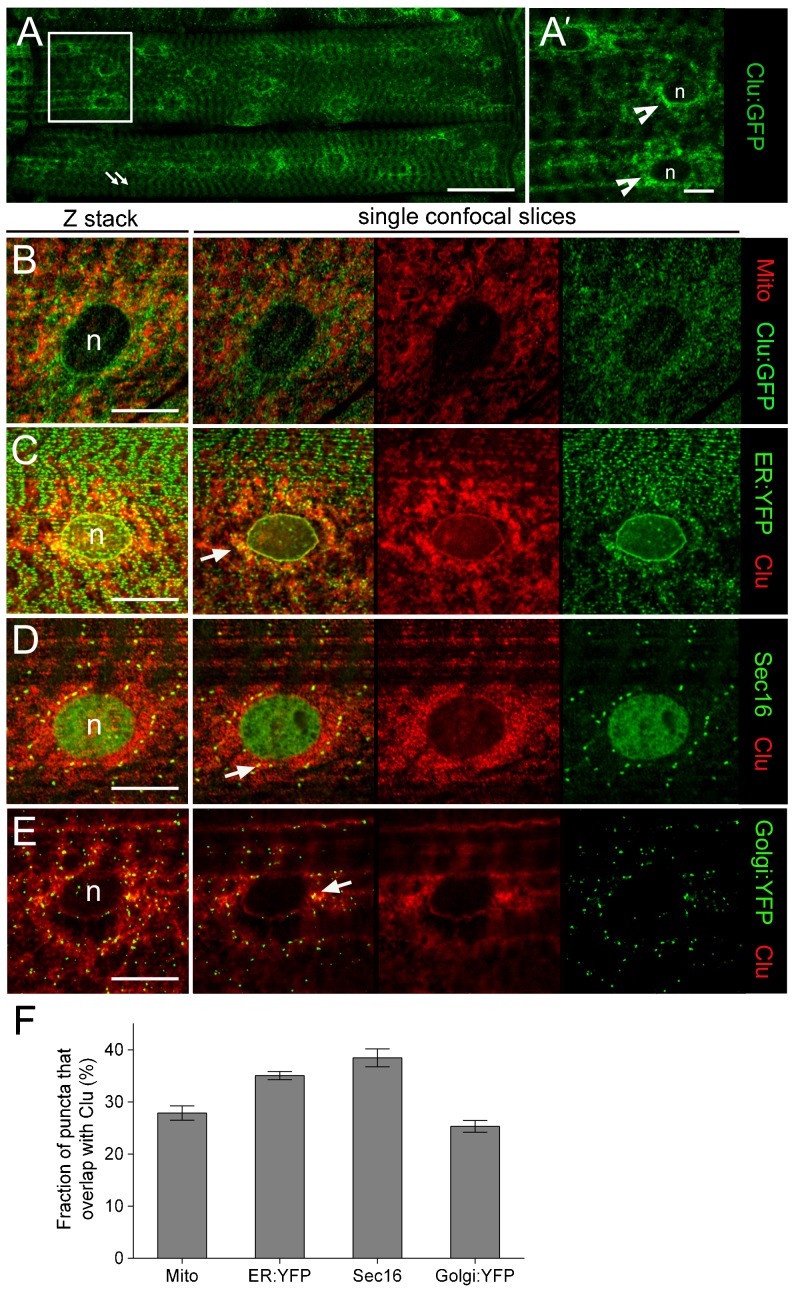
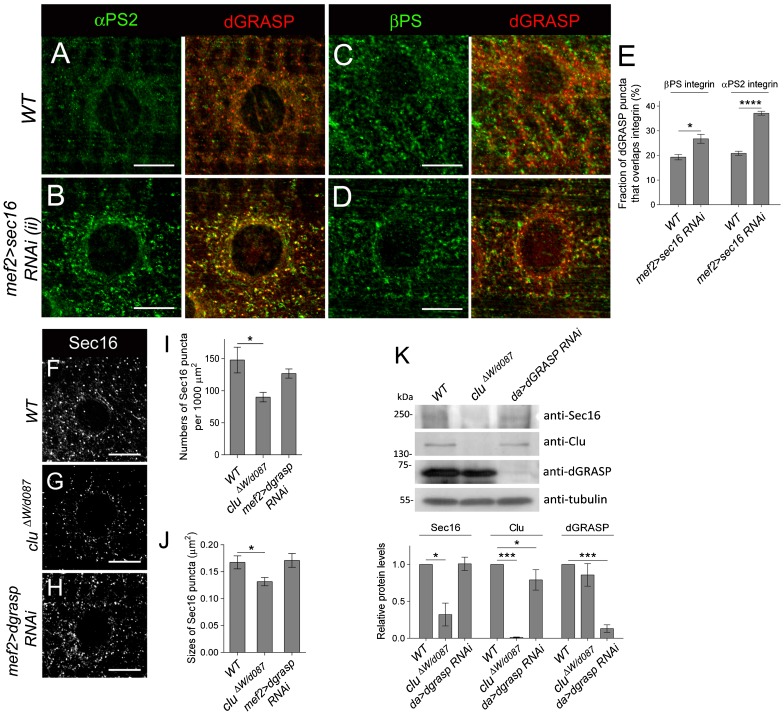
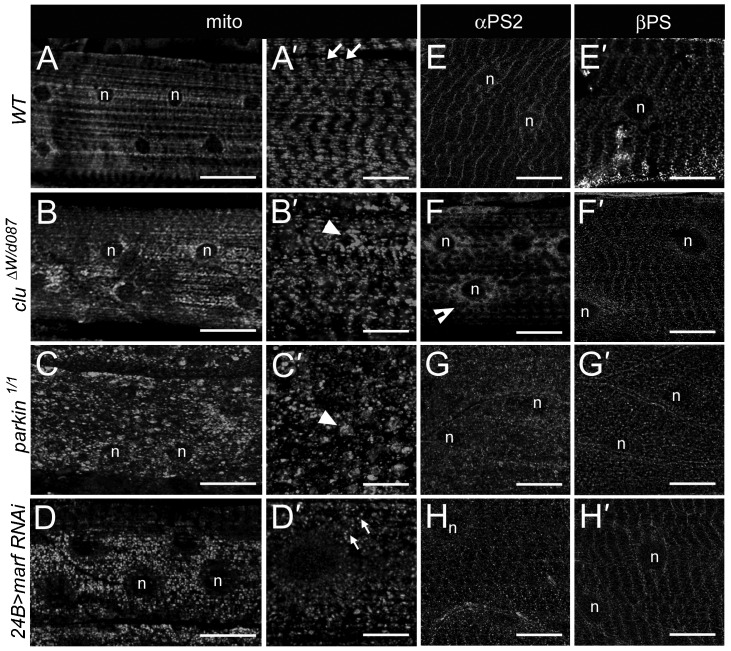
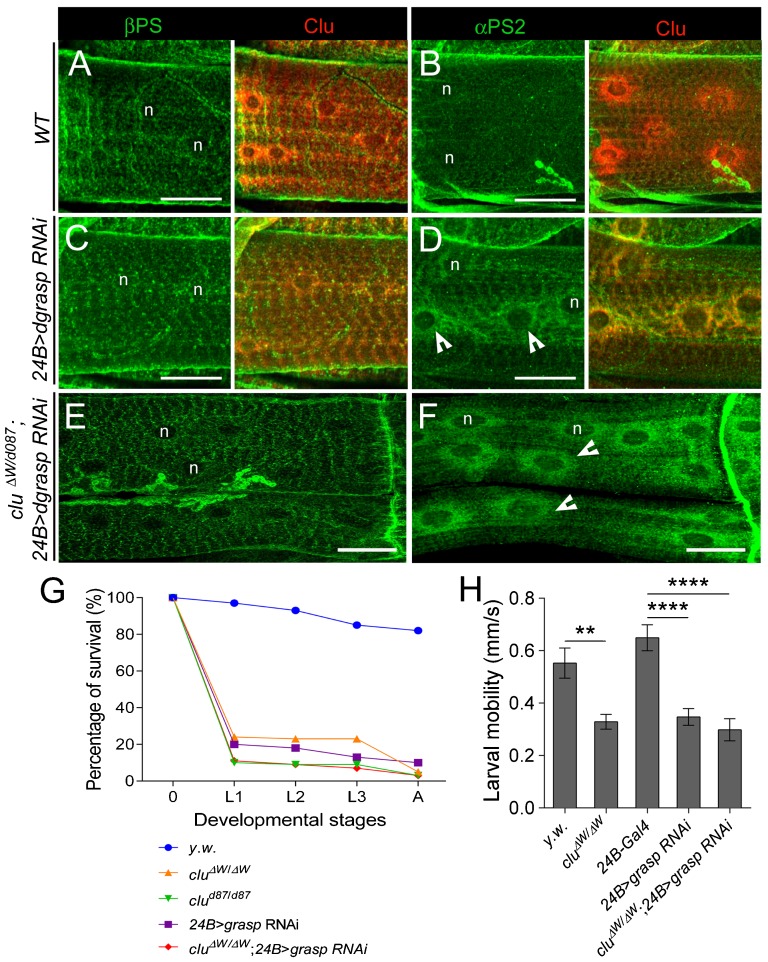
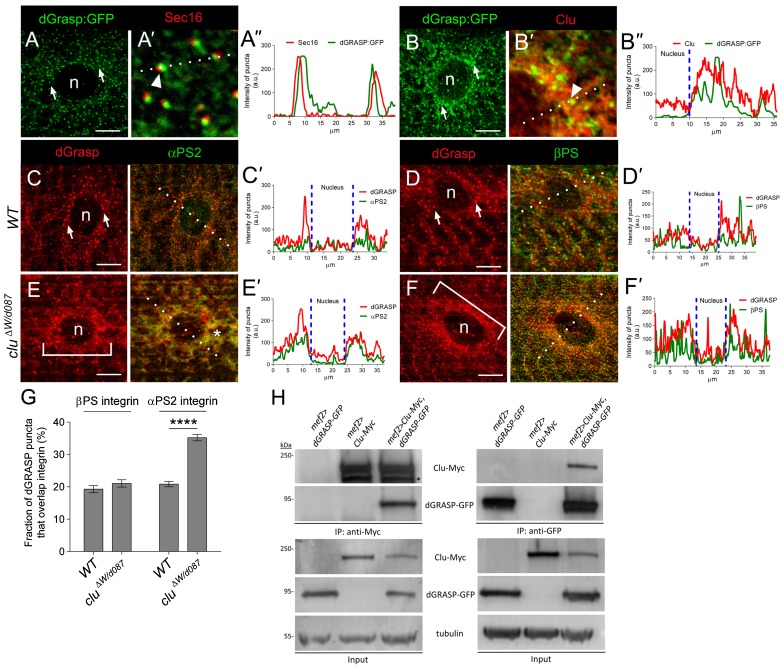
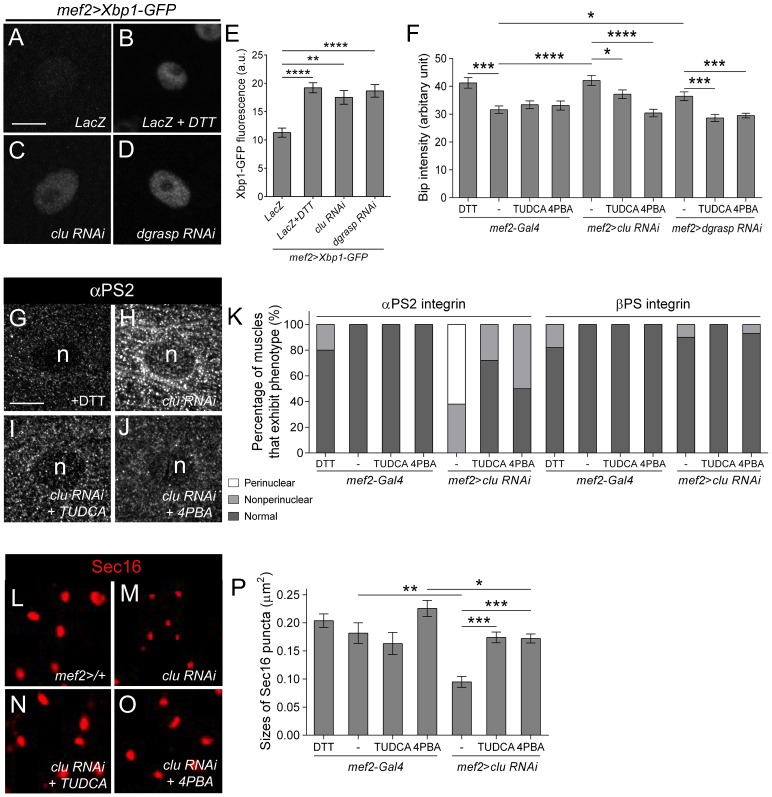
Drosophila Clueless (Clu) and its conserved orthologs are known for their role in the prevention of mitochondrial clustering. Here, we uncover a new role for Clu in the delivery of integrin subunits in muscle tissue. In clu mutants, αPS2 integrin, but not βPS integrin, abnormally accumulates in a perinuclear endoplasmic reticulum (ER) subdomain, a site that mirrors the endogenous localization of Clu. Loss of components essential for mitochondrial distribution do not phenocopy the clu mutant αPS2 phenotype. Conversely, RNAi knockdown of the Drosophila Golgi reassembly and stacking protein GRASP55/65 (dGRASP) recapitulates clu defects, including the abnormal accumulation of αPS2 and larval locomotor activity. Both Clu and dGRASP proteins physically interact and loss of Clu displaces dGRASP from ER exit sites, suggesting that Clu cooperates with dGRASP for the exit of αPS2 from a perinuclear subdomain in the ER. We also found that Clu and dGRASP loss of function leads to ER stress and that the stability of the ER exit site protein Sec16 is severely compromised in the clu mutants, thus explaining the ER accumulation of αPS2. Remarkably, exposure of clu RNAi larvae to chemical chaperones restores both αPS2 delivery and functional ER exit sites. We propose that Clu together with dGRASP prevents ER stress and therefore maintains Sec16 stability essential for the functional organization of perinuclear early secretory pathway. This, in turn, is essential for integrin subunit αPS2 ER exit in Drosophila larval myofibers.
Keywords: Clueless; Drosophila; Integrin; Muscle; Trafficking; dGRASP.
© 2015. Published by The Company of Biologists Ltd.
Conflict of interest statement
Figures
Similar articles
-
dGRASP localization and function in the early exocytic pathway in Drosophila S2 cells.Mol Biol Cell. 2005 Sep;16(9):4061-72. doi: 10.1091/mbc.e04-10-0938. Epub 2005 Jun 22. Mol Biol Cell. 2005. PMID: 15975913 Free PMC article.
-
Drosophila clueless is highly expressed in larval neuroblasts, affects mitochondrial localization and suppresses mitochondrial oxidative damage.PLoS One. 2013;8(1):e54283. doi: 10.1371/journal.pone.0054283. Epub 2013 Jan 16. PLoS One. 2013. PMID: 23342118 Free PMC article.
-
dGRASP-mediated noncanonical integrin secretion is required for Drosophila epithelial remodeling.Dev Cell. 2008 Feb;14(2):171-82. doi: 10.1016/j.devcel.2007.12.006. Dev Cell. 2008. PMID: 18267086
-
Sec16 in conventional and unconventional exocytosis: Working at the interface of membrane traffic and secretory autophagy?J Cell Physiol. 2017 Dec;232(12):3234-3243. doi: 10.1002/jcp.25842. Epub 2017 Apr 27. J Cell Physiol. 2017. PMID: 28160489 Review.
-
Cellular responses to endoplasmic reticulum stress and apoptosis.Apoptosis. 2009 Aug;14(8):996-1007. doi: 10.1007/s10495-009-0341-y. Epub 2009 Apr 10. Apoptosis. 2009. PMID: 19360473 Review.
Cited by
-
Clueless forms dynamic, insulin-responsive bliss particles sensitive to stress.Dev Biol. 2020 Mar 15;459(2):149-160. doi: 10.1016/j.ydbio.2019.12.004. Epub 2019 Dec 16. Dev Biol. 2020. PMID: 31837288 Free PMC article.
-
Drosophila clueless is involved in Parkin-dependent mitophagy by promoting VCP-mediated Marf degradation.Hum Mol Genet. 2016 May 15;25(10):1946-1964. doi: 10.1093/hmg/ddw067. Epub 2016 Feb 29. Hum Mol Genet. 2016. PMID: 26931463 Free PMC article.
-
Drosophila Clueless ribonucleoprotein particles display novel dynamics that rely on the availability of functional protein and polysome equilibrium.bioRxiv [Preprint]. 2024 Aug 22:2024.08.21.609023. doi: 10.1101/2024.08.21.609023. bioRxiv. 2024. Update in: J Cell Sci. 2025 May 1;138(9):jcs263730. doi: 10.1242/jcs.263730. PMID: 39229069 Free PMC article. Updated. Preprint.
-
Sec16A is critical for both conventional and unconventional secretion of CFTR.Sci Rep. 2017 Jan 9;7:39887. doi: 10.1038/srep39887. Sci Rep. 2017. PMID: 28067262 Free PMC article.
-
Investigating the cellular functions of β-Glucosidases for synthesis of lignocellulose-degrading enzymes in Trichoderma reesei.Eng Microbiol. 2023 Jul 5;3(4):100105. doi: 10.1016/j.engmic.2023.100105. eCollection 2023 Dec. Eng Microbiol. 2023. PMID: 39628917 Free PMC article.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
