

Targeting the spliceosome in chronic lymphocytic leukemia with the macrolides FD-895 and pladienolide-B
- PMID: 25862704
- PMCID: PMC4486229
- DOI: 10.3324/haematol.2014.122069
Targeting the spliceosome in chronic lymphocytic leukemia with the macrolides FD-895 and pladienolide-B
Abstract
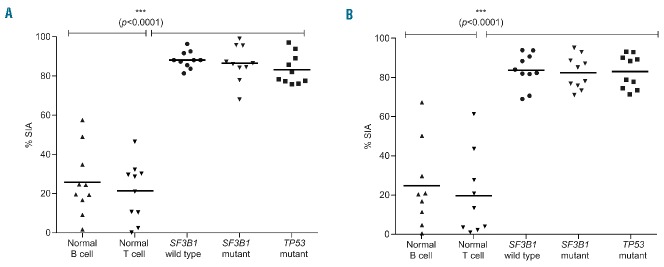
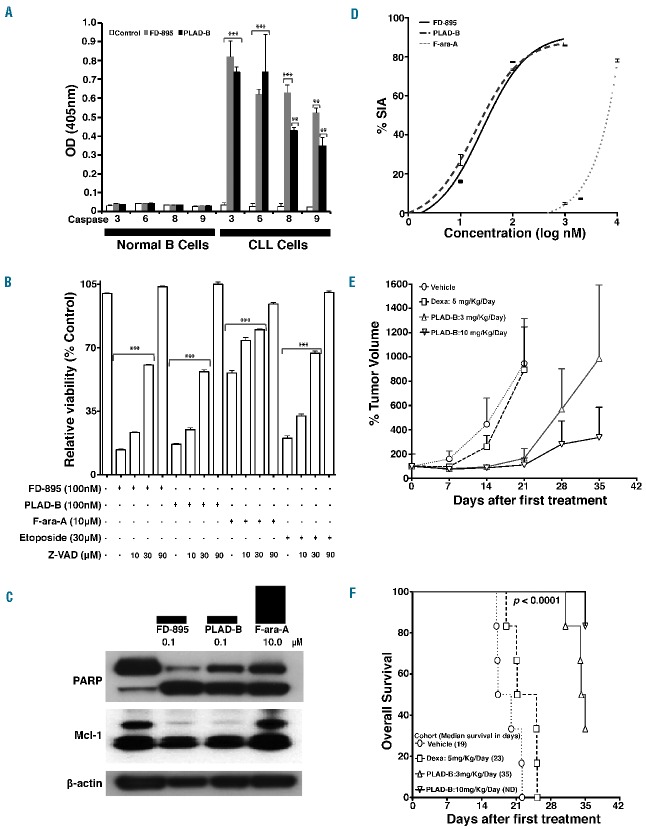
RNA splicing plays a fundamental role in human biology. Its relevance in cancer is rapidly emerging as demonstrated by spliceosome mutations that determine the prognosis of patients with hematologic malignancies. We report studies using FD-895 and pladienolide-B in primary leukemia cells derived from patients with chronic lymphocytic leukemia and leukemia-lymphoma cell lines. We found that FD-895 and pladienolide-B induce an early pattern of mRNA intron retention - spliceosome modulation. This process was associated with apoptosis preferentially in cancer cells as compared to normal lymphocytes. The pro-apoptotic activity of these compounds was observed regardless of poor prognostic factors such as Del(17p), TP53 or SF3B1 mutations and was able to overcome the protective effect of culture conditions that resemble the tumor microenvironment. In addition, the activity of these compounds was observed not only in vitro but also in vivo using the A20 lymphoma murine model. Overall, these findings give evidence for the first time that spliceosome modulation is a valid target in chronic lymphocytic leukemia and provide an additional rationale for the development of spliceosome modulators for cancer therapy.
Copyright© Ferrata Storti Foundation.
Figures
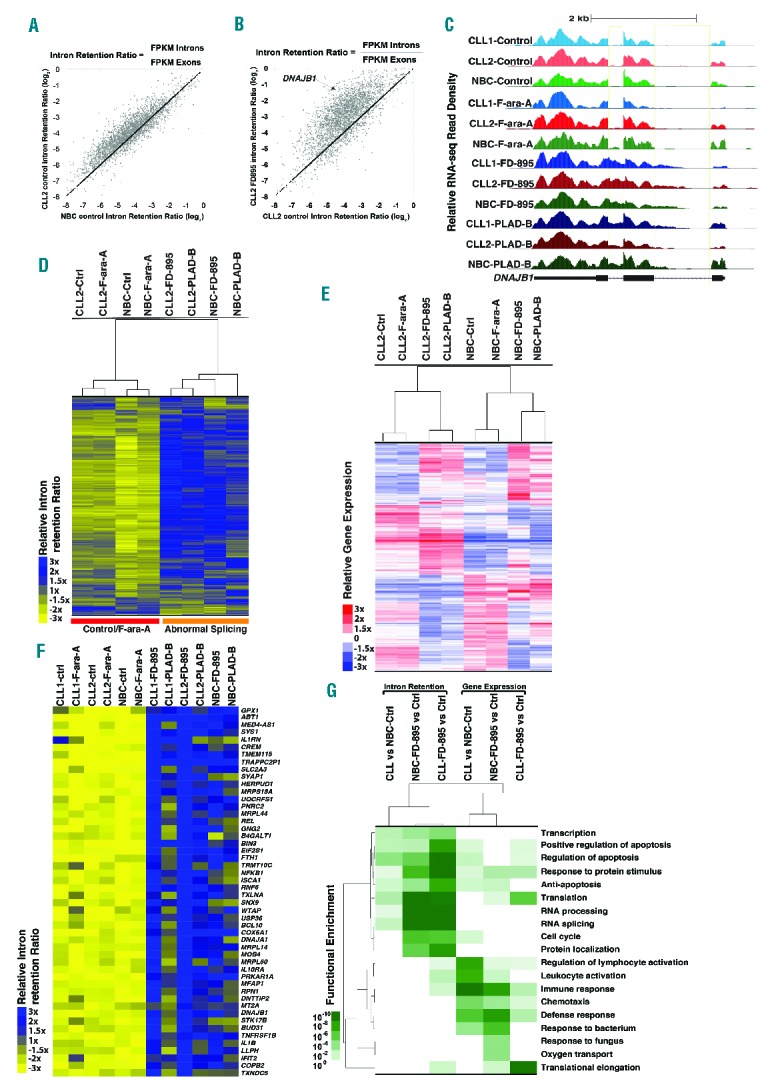
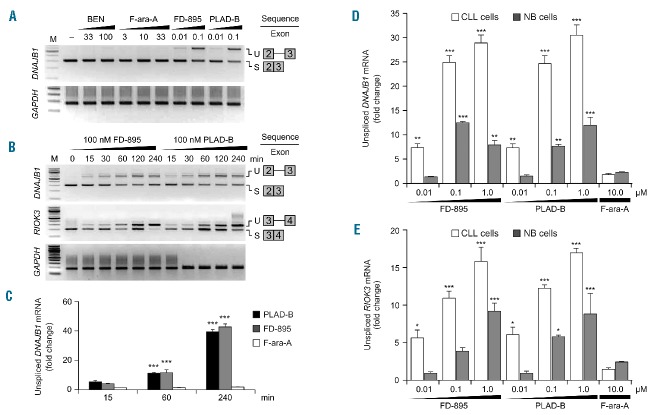
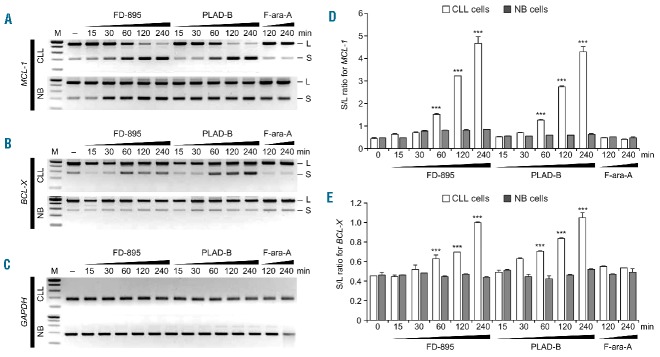
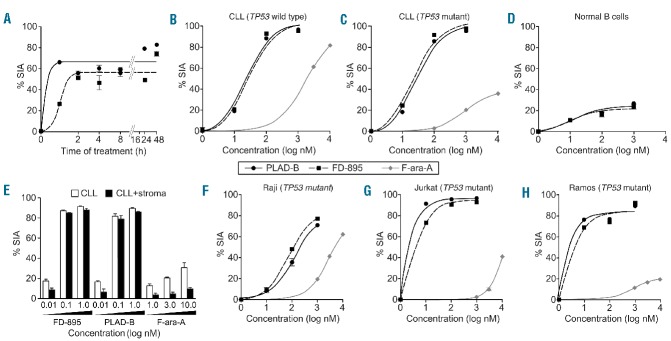
References
-
- Rai KR. Pathophysiologic mechanisms of chronic lymphocytic leukemia and their application to therapy. Exp Hematol. 2007;35(4 Suppl 1):134–136. - PubMed
-
- Hallek M, Fischer K, Fingerle-Rowson G, et al. Addition of rituximab to fludarabine and cyclophosphamide in patients with chronic lymphocytic leukaemia: a randomised, open-label, phase 3 trial. Lancet. 2010;376(9747):1164–1174. - PubMed
-
- Castro JE. Treatment of patients with chronic lymphocytic leukemia with 17p deletion: the saga continues. Leuk Lymphoma. 2012;53(2):179–180. - PubMed
-
- Hallek M. Signaling the end of chronic lymphocytic leukemia: new frontline treatment strategies. Blood. 2013;122(23):3723–3734. - PubMed
-
- Kornblihtt AR, Schor IE, Alló M, Dujardin G, Petrillo E, Muñoz MJ. Alternative splicing: a pivotal step between eukaryotic transcription and translation. Nat Rev Mol Cell Biol. 2013;14(3):153–165. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
