

Redox theory of aging
- PMID: 25863726
- PMCID: PMC4392062
- DOI: 10.1016/j.redox.2015.03.004
Redox theory of aging
Abstract
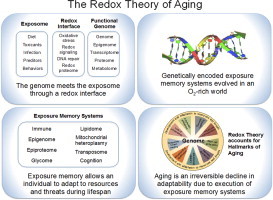
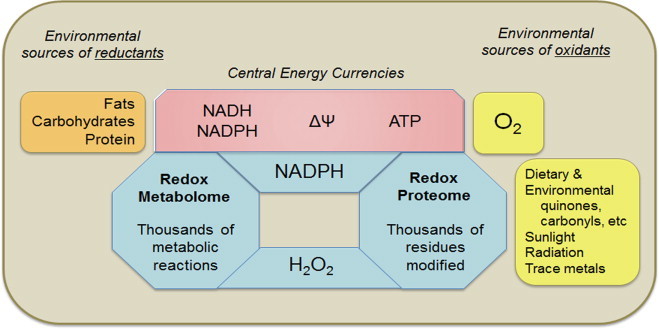
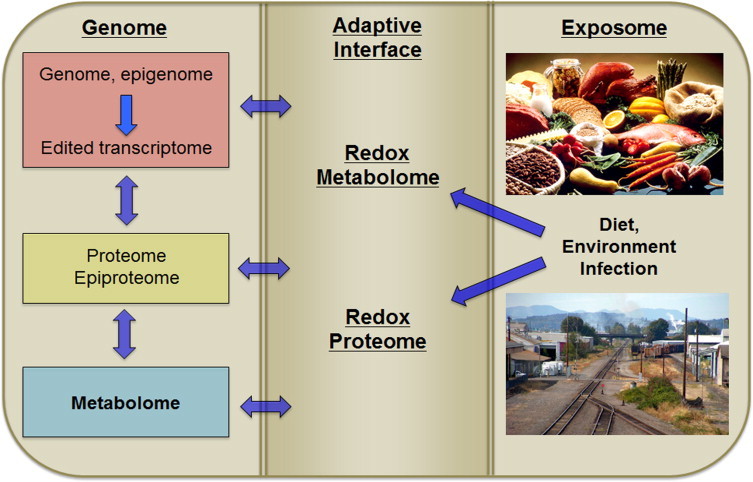
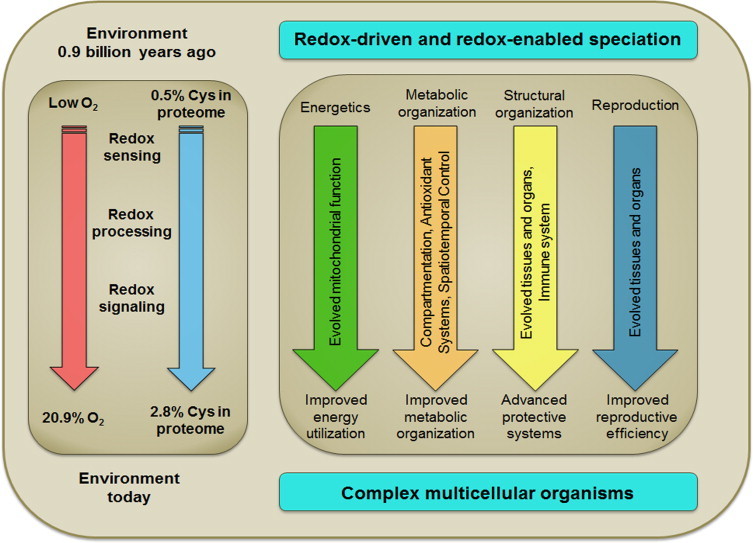
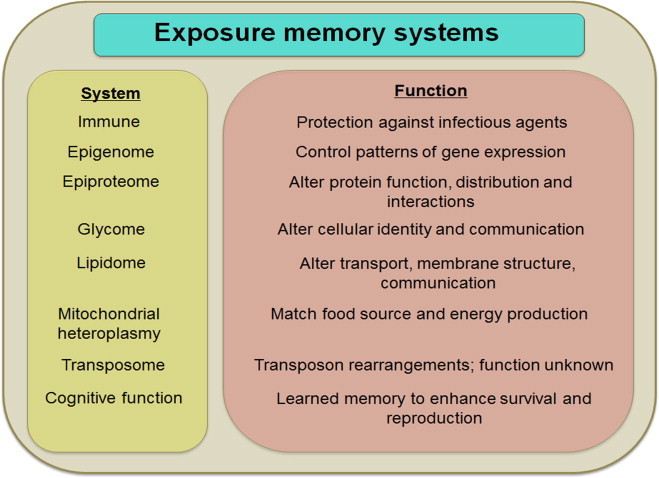
Metazoan genomes encode exposure memory systems to enhance survival and reproductive potential by providing mechanisms for an individual to adjust during lifespan to environmental resources and challenges. These systems are inherently redox networks, arising during evolution of complex systems with O2 as a major determinant of bioenergetics, metabolic and structural organization, defense, and reproduction. The network structure decreases flexibility from conception onward due to differentiation and cumulative responses to environment (exposome). The redox theory of aging is that aging is a decline in plasticity of genome-exposome interaction that occurs as a consequence of execution of differentiation and exposure memory systems. This includes compromised mitochondrial and bioenergetic flexibility, impaired food utilization and metabolic homeostasis, decreased barrier and defense capabilities and loss of reproductive fidelity and fecundity. This theory accounts for hallmarks of aging, including failure to maintain oxidative or xenobiotic defenses, mitochondrial integrity, proteostasis, barrier structures, DNA repair, telomeres, immune function, metabolic regulation and regenerative capacity.
Keywords: Oxidative stress; Redox signaling; Redox systems biology.
Copyright © 2015. Published by Elsevier B.V.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
