

Interictal spike frequency varies with ovarian cycle stage in a rat model of epilepsy
- PMID: 25864929
- PMCID: PMC4446145
- DOI: 10.1016/j.expneurol.2015.04.003
Interictal spike frequency varies with ovarian cycle stage in a rat model of epilepsy
Abstract
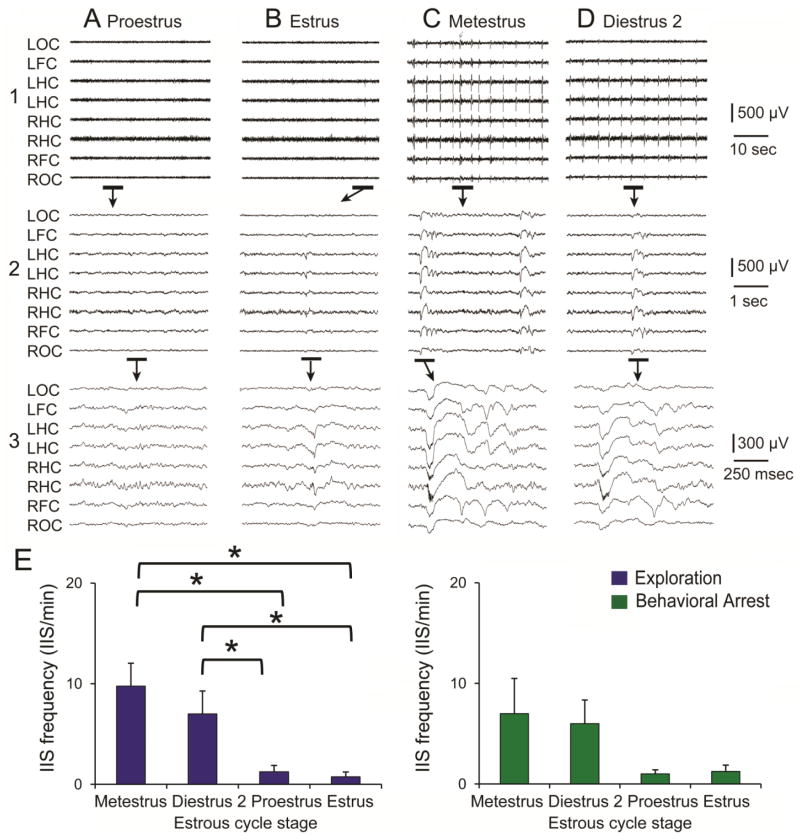
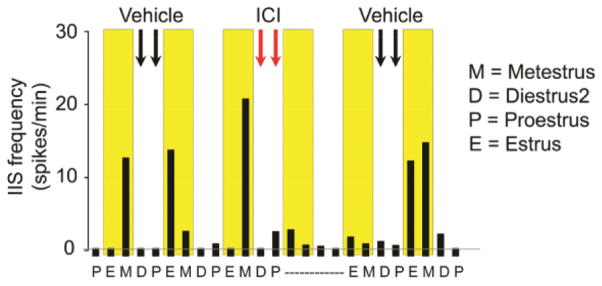
In catamenial epilepsy, seizures exhibit a cyclic pattern that parallels the menstrual cycle. Many studies suggest that catamenial seizures are caused by fluctuations in gonadal hormones during the menstrual cycle, but this has been difficult to study in rodent models of epilepsy because the ovarian cycle in rodents, called the estrous cycle, is disrupted by severe seizures. Thus, when epilepsy is severe, estrous cycles become irregular or stop. Therefore, we modified kainic acid (KA)- and pilocarpine-induced status epilepticus (SE) models of epilepsy so that seizures were rare for the first months after SE, and conducted video-EEG during this time. The results showed that interictal spikes (IIS) occurred intermittently. All rats with regular 4-day estrous cycles had IIS that waxed and waned with the estrous cycle. The association between the estrous cycle and IIS was strong: if the estrous cycles became irregular transiently, IIS frequency also became irregular, and when the estrous cycle resumed its 4-day pattern, IIS frequency did also. Furthermore, when rats were ovariectomized, or males were recorded, IIS frequency did not show a 4-day pattern. Systemic administration of an estrogen receptor antagonist stopped the estrous cycle transiently, accompanied by transient irregularity of the IIS pattern. Eventually all animals developed severe, frequent seizures and at that time both the estrous cycle and the IIS became irregular. We conclude that the estrous cycle entrains IIS in the modified KA and pilocarpine SE models of epilepsy. The data suggest that the ovarian cycle influences more aspects of epilepsy than seizure susceptibility.
Keywords: Animal model; Epilepsy; Hormone; Kainic acid; Neuropathology; Seizure; Women.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures








References
-
- Penovich PE, Helmers S. Catamenial epilepsy. Int Rev Neurobiol. 2008;83:79–90. - PubMed
-
- Verrotti A, Laus M, Coppola G, Parisi P, Mohn A, Chiarelli F. Catamenial epilepsy: Hormonal aspects. Gynecol Endocrinol. 2010;26:783–790. - PubMed
-
- Herzog AG. Catamenial epilepsy: Definition, prevalence pathophysiology and treatment. Seizure. 2008;17:151–159. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
