

Sustained activation of GABAA receptors in the suprachiasmatic nucleus mediates light-induced phase delays of the circadian clock: a novel function of ionotropic receptors
- PMID: 25865743
- PMCID: PMC4510013
- DOI: 10.1111/ejn.12918
Sustained activation of GABAA receptors in the suprachiasmatic nucleus mediates light-induced phase delays of the circadian clock: a novel function of ionotropic receptors
Abstract
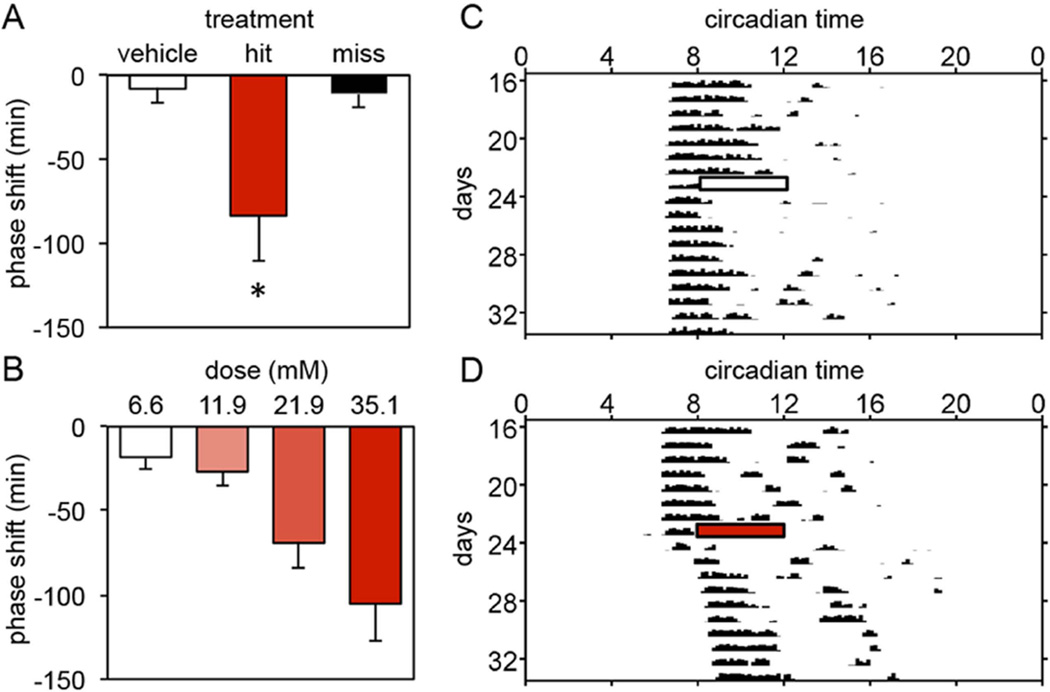
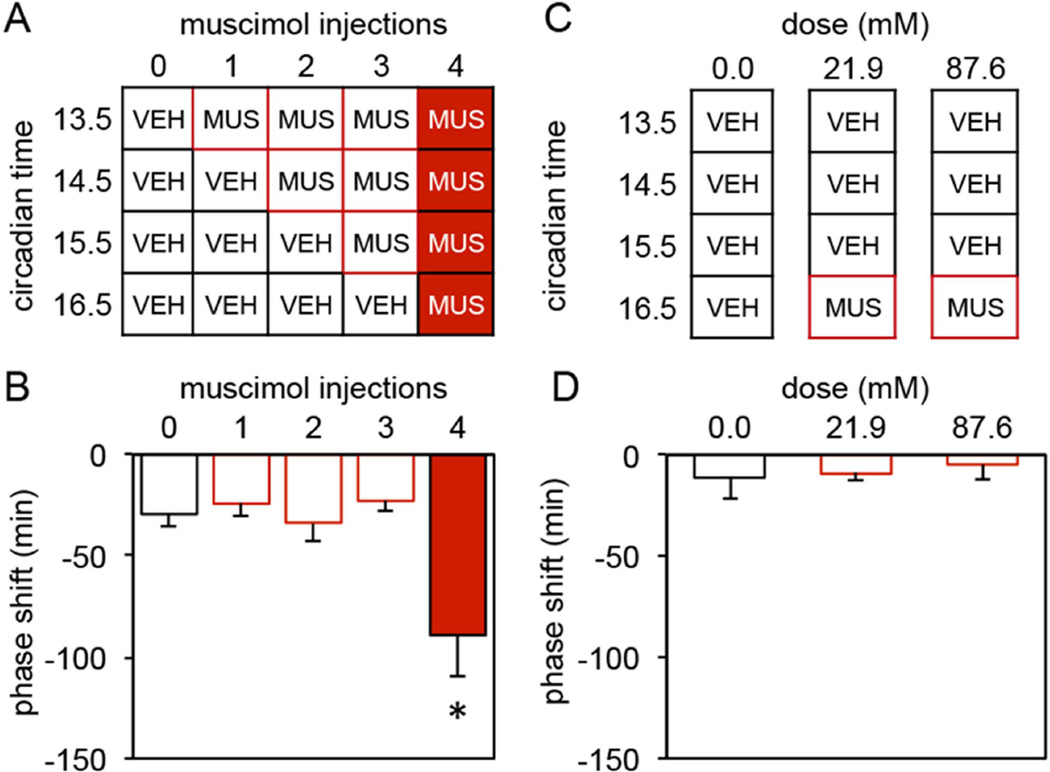
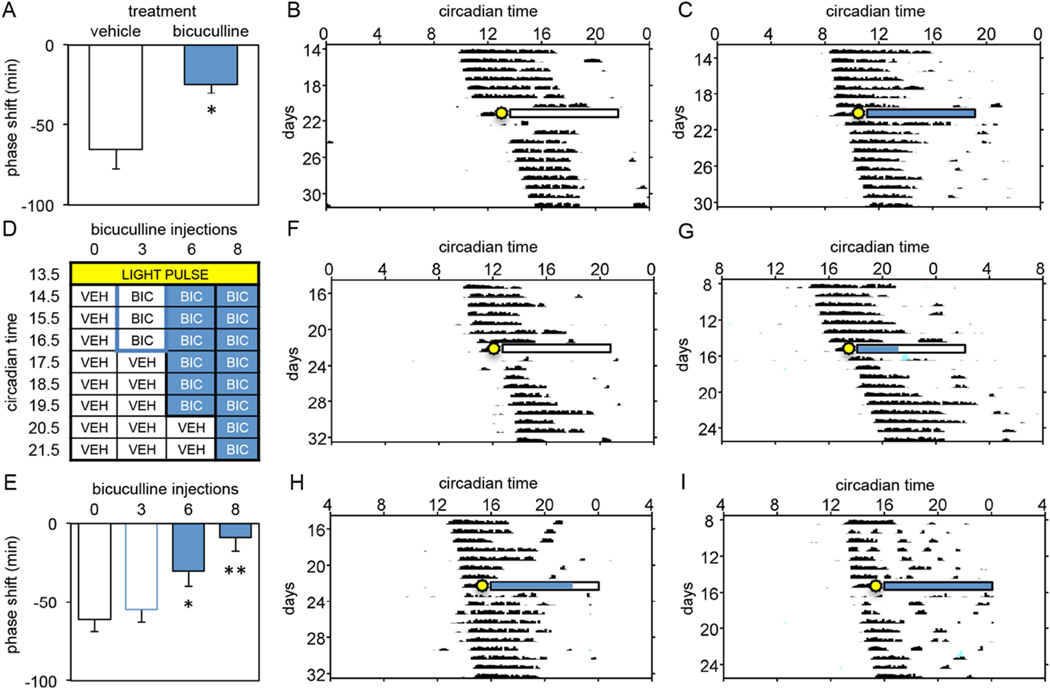
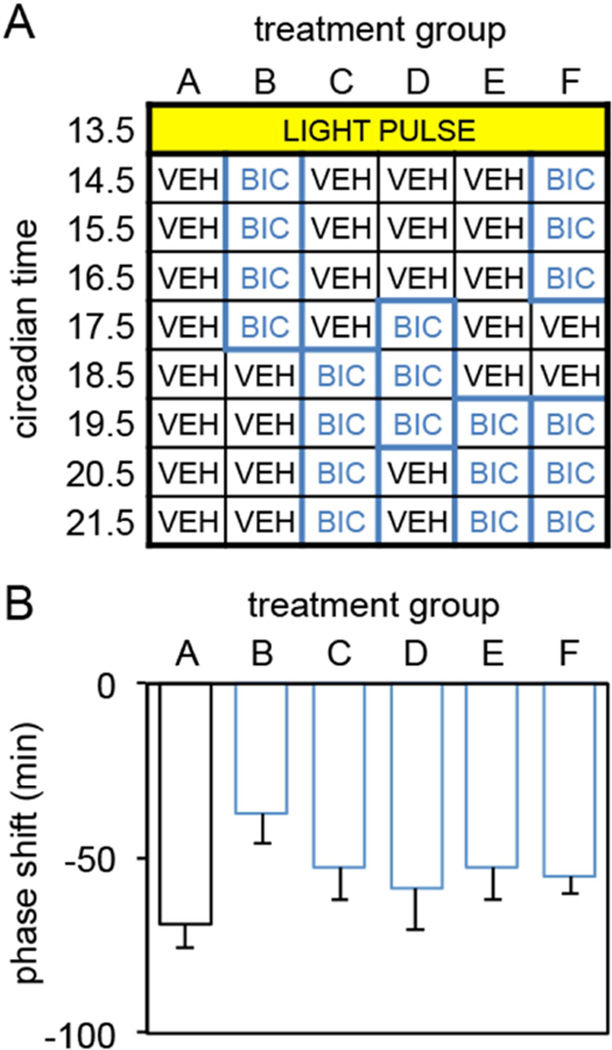
The suprachiasmatic nucleus (SCN) contains a circadian clock that generates endogenous rhythmicity and entrains that rhythmicity with the day-night cycle. The neurochemical events that transduce photic input within the SCN and mediate entrainment by resetting the molecular clock have yet to be defined. Because GABA is contained in nearly all SCN neurons we tested the hypothesis that GABA serves as this signal in studies employing Syrian hamsters (Mesocricetus auratus). Activation of GABAA receptors was found to be necessary and sufficient for light to induce phase delays of the clock. Remarkably, the sustained activation of GABAA receptors for more than three consecutive hours was necessary to phase-delay the clock. The duration of GABAA receptor activation required to induce phase delays would not have been predicted by either the prevalent theory of circadian entrainment or by expectations regarding the duration of ionotropic receptor activation necessary to produce functional responses. Taken together, these data identify a novel neurochemical mechanism essential for phase-delaying the 'master' circadian clock within the SCN as well as identifying an unprecedented action of an amino acid neurotransmitter involving the sustained activation of ionotropic receptors.
Keywords: entrainment; hamster; neurotransmission; pacemaker; photic.
© 2015 Federation of European Neuroscience Societies and John Wiley & Sons Ltd.
Figures
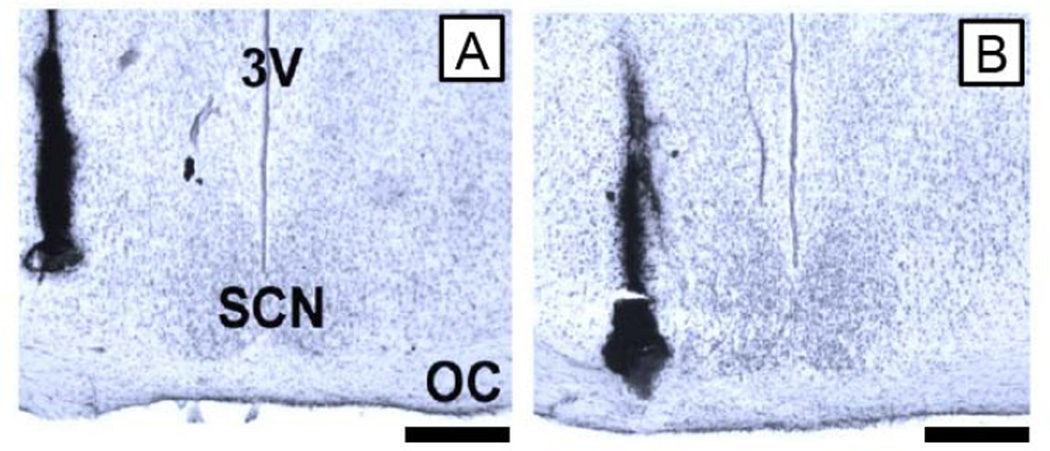
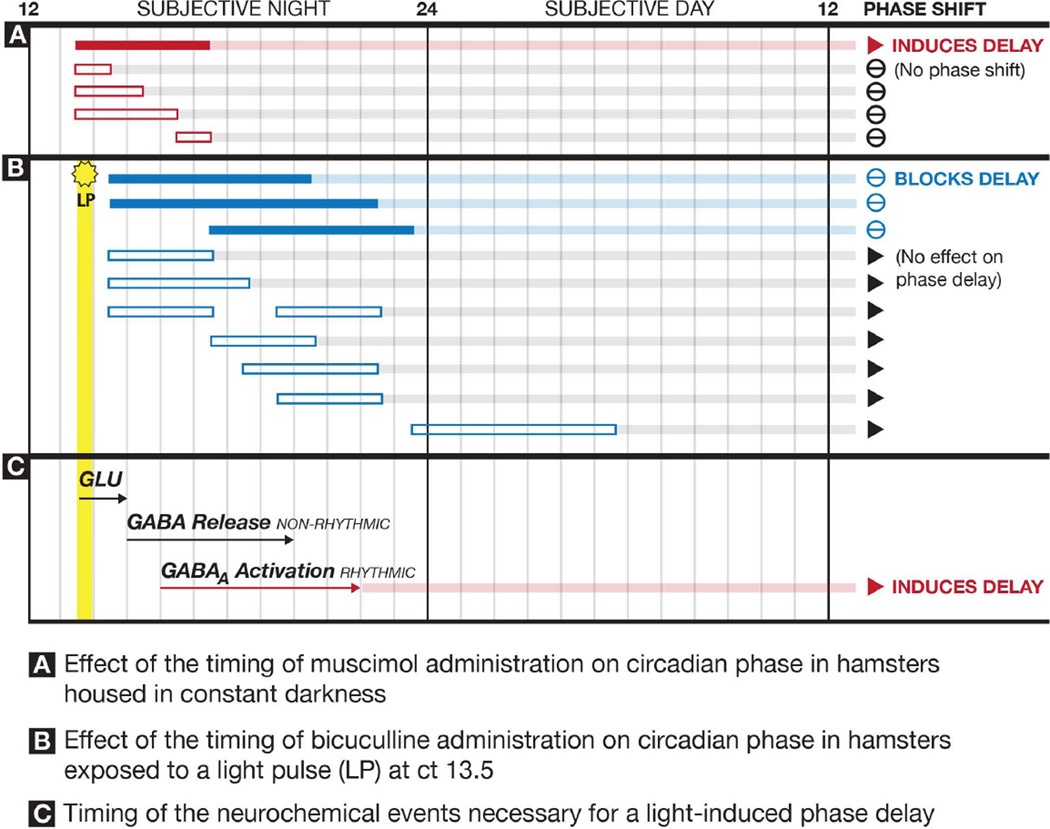
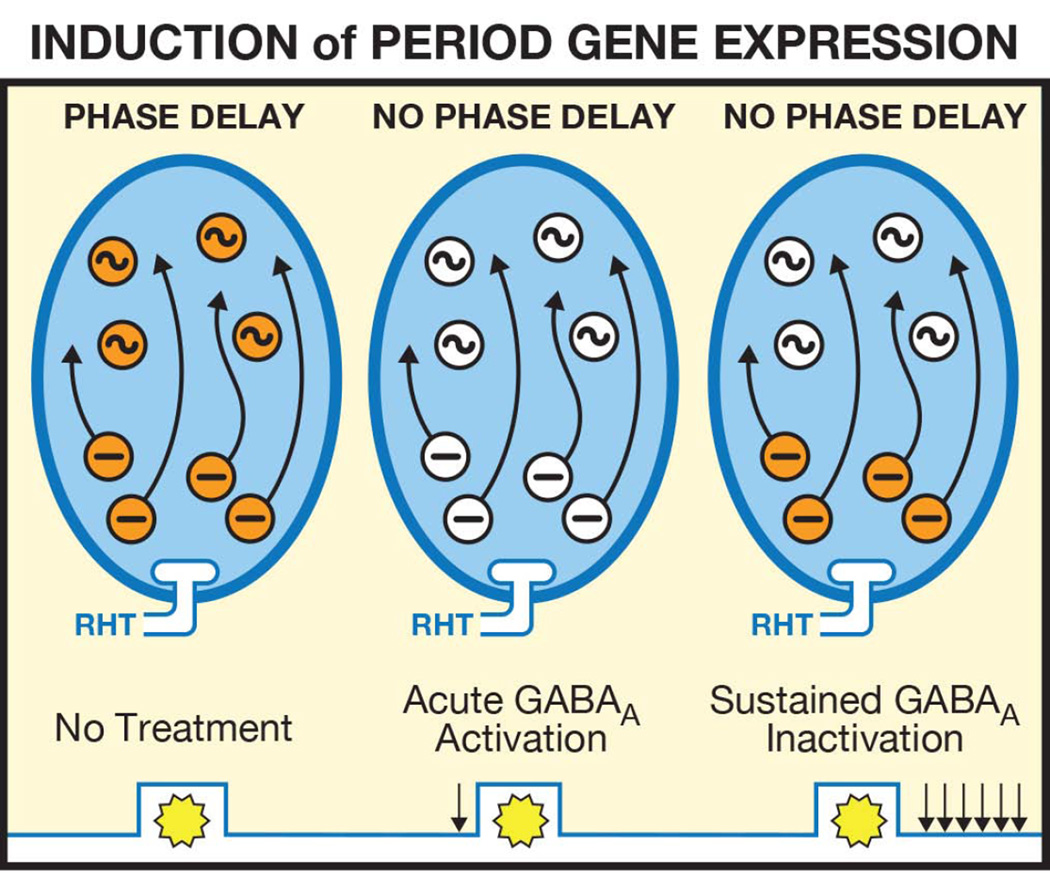
Similar articles
-
GABA interacts with photic signaling in the suprachiasmatic nucleus to regulate circadian phase shifts.Neuroscience. 2002;109(4):773-8. doi: 10.1016/s0306-4522(01)00519-x. Neuroscience. 2002. PMID: 11927159
-
Interactions of GABA A receptor activation and light on period mRNA expression in the suprachiasmatic nucleus.J Biol Rhythms. 2008 Feb;23(1):16-25. doi: 10.1177/0748730407310785. J Biol Rhythms. 2008. PMID: 18258754
-
GABAA receptor activation suppresses Period 1 mRNA and Period 2 mRNA in the suprachiasmatic nucleus during the mid-subjective day.Eur J Neurosci. 2006 Jun;23(12):3328-36. doi: 10.1111/j.1460-9568.2006.04857.x. Eur J Neurosci. 2006. PMID: 16820022
-
The cholinergic system, circadian rhythmicity, and time memory.Behav Brain Res. 2011 Aug 10;221(2):466-80. doi: 10.1016/j.bbr.2010.11.039. Epub 2010 Nov 27. Behav Brain Res. 2011. PMID: 21115064 Review.
-
Assessing ethanol's actions in the suprachiasmatic circadian clock using in vivo and in vitro approaches.Alcohol. 2015 Jun;49(4):321-339. doi: 10.1016/j.alcohol.2014.07.016. Epub 2014 Oct 18. Alcohol. 2015. PMID: 25457753 Free PMC article. Review.
Cited by
-
Systematic review of drugs that modify the circadian system's phase-shifting responses to light exposure.Neuropsychopharmacology. 2022 Mar;47(4):866-879. doi: 10.1038/s41386-021-01251-8. Epub 2021 Dec 27. Neuropsychopharmacology. 2022. PMID: 34961774 Free PMC article.
-
A Symphony of Signals: Intercellular and Intracellular Signaling Mechanisms Underlying Circadian Timekeeping in Mice and Flies.Int J Mol Sci. 2019 May 13;20(9):2363. doi: 10.3390/ijms20092363. Int J Mol Sci. 2019. PMID: 31086044 Free PMC article. Review.
-
Associations between perioperative sleep patterns and clinical outcomes in patients with intracranial tumors: a correlation study.Front Neurol. 2023 Sep 5;14:1242360. doi: 10.3389/fneur.2023.1242360. eCollection 2023. Front Neurol. 2023. PMID: 37731854 Free PMC article.
-
Retina-attached slice recording reveals light-triggered tonic GABA signaling in suprachiasmatic nucleus.Mol Brain. 2021 Nov 27;14(1):171. doi: 10.1186/s13041-021-00881-9. Mol Brain. 2021. PMID: 34838118 Free PMC article.
-
Temporal Regulation of GABAA Receptor Subunit Expression: Role in Synaptic and Extrasynaptic Communication in the Suprachiasmatic Nucleus.eNeuro. 2017 May 1;4(2):ENEURO.0352-16.2017. doi: 10.1523/ENEURO.0352-16.2017. eCollection 2017 Mar-Apr. eNeuro. 2017. PMID: 28466071 Free PMC article.
References
-
- Albus H, Vansteensel MJ, Michel S, Block GD, Meijer JH. A GABAergic mechanism is necessary for coupling dissociable ventral and dorsal regional oscillators within the circadian clock. Curr Biol. 2005;15:886–893. - PubMed
-
- Antle MC, Smith VM, Sterniczuk R, Yamakawa GR, Rakai BD. Physiological responses of the circadian clock to acute light exposure at night. Rev Endocr Metab Disord. 2009;10:279–291. - PubMed
-
- Cohen R, Albers HE. Disruption of human circadian and cognitive regulation following a discrete hypothalamic lesion: A case study. Neurol. 1991;41:726–729. - PubMed
-
- Colwell CS, Menaker M. Regulation of circadian rhythms by excitatory amino acids. In: Brann DW, Mahesh VB, editors. Excitatory amino acids: Their role in neuroendocrine function. New York: CRC Press; 1995. pp. 223–252.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
