

The Mechanochemical Cycle of Mammalian Kinesin-2 KIF3A/B under Load
- PMID: 25866395
- PMCID: PMC4422762
- DOI: 10.1016/j.cub.2015.03.013
The Mechanochemical Cycle of Mammalian Kinesin-2 KIF3A/B under Load
Abstract
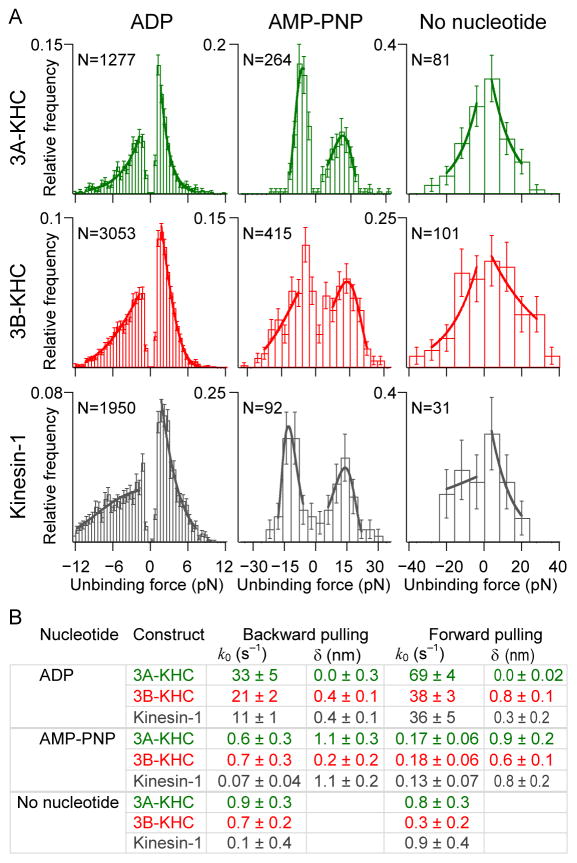
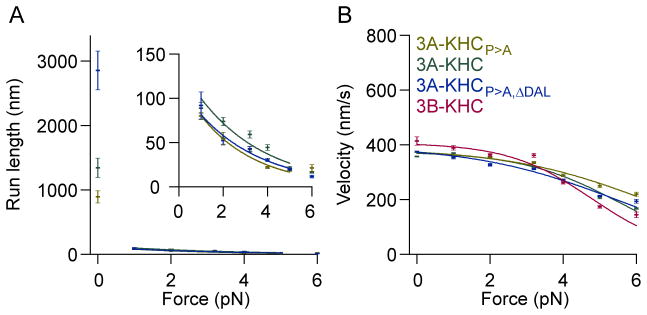
The response of motor proteins to external loads underlies their ability to work in teams and determines the net speed and directionality of cargo transport. The mammalian kinesin-2, KIF3A/B, is a heterotrimeric motor involved in intraflagellar transport and vesicle motility in neurons. Bidirectional cargo transport is known to result from the opposing activities of KIF3A/B and dynein bound to the same cargo, but the load-dependent properties of kinesin-2 are poorly understood. We used a feedback-controlled optical trap to probe the velocity, run length, and unbinding kinetics of mouse KIF3A/B under various loads and nucleotide conditions. The kinesin-2 motor velocity is less sensitive than kinesin-1 to external forces, but its processivity diminishes steeply with load, and the motor was observed occasionally to slip and reattach. Each motor domain was characterized by studying homodimeric constructs, and a global fit to the data resulted in a comprehensive pathway that quantifies the principal force-dependent kinetic transitions. The properties of the KIF3A/B heterodimer are intermediate between the two homodimers, and the distinct load-dependent behavior is attributable to the properties of the motor domains and not to the neck linkers or the coiled-coil stalk. We conclude that the force-dependent movement of KIF3A/B differs significantly from conventional kinesin-1. Against opposing dynein forces, KIF3A/B motors are predicted to rapidly unbind and rebind, resulting in qualitatively different transport behavior from kinesin-1.
Copyright © 2015 Elsevier Ltd. All rights reserved.
Figures
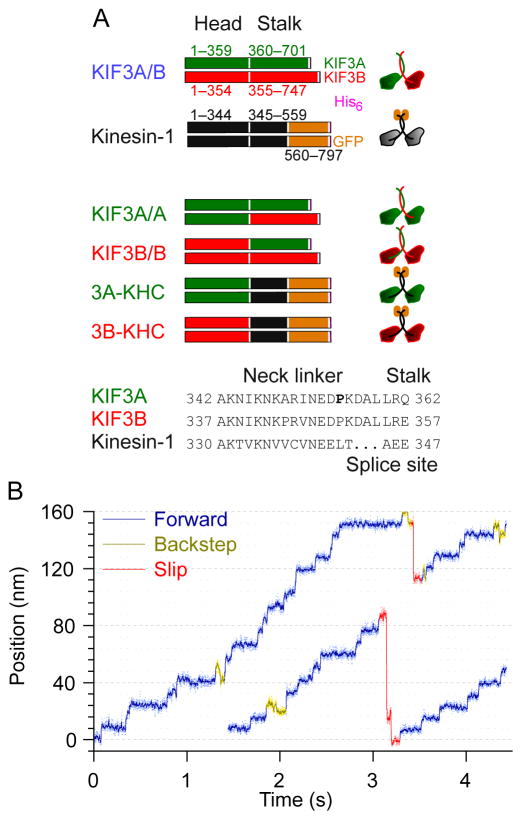
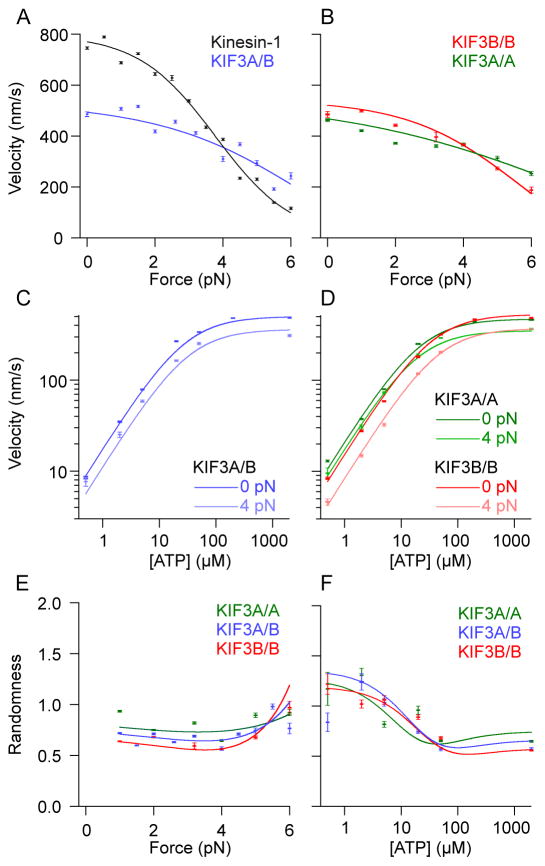
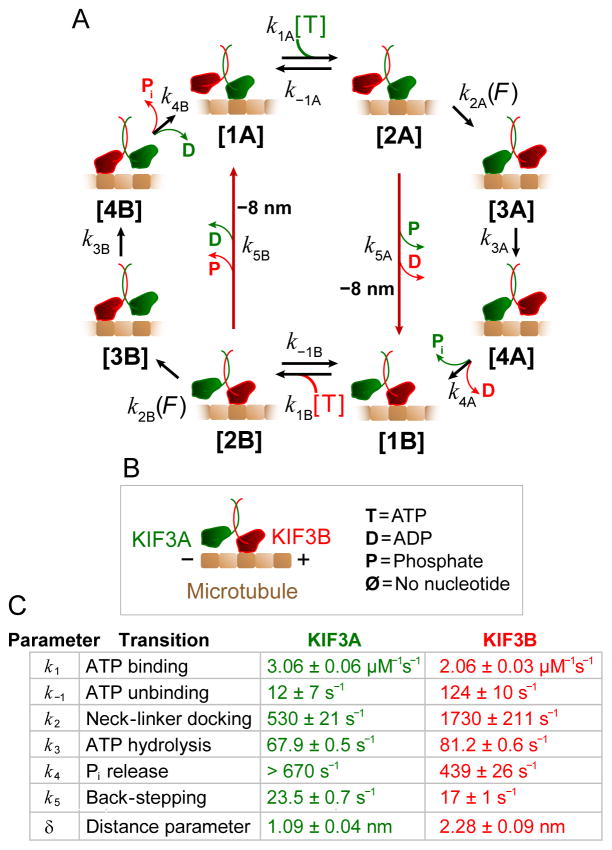
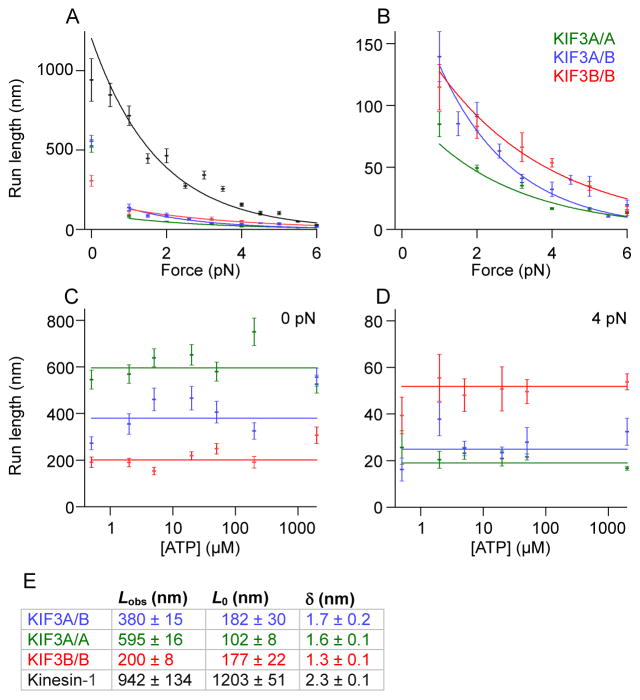
References
-
- Haraguchi K, Hayashi T, Jimbo T, Yamamoto T, Akiyama T. Role of the kinesin-2 family protein, KIF3, during mitosis. J Biol Chem. 2006;281:4094–4099. - PubMed
-
- Hirokawa N, Noda Y. Intracellular transport and kinesin superfamily proteins, KIFs: structure, function, and dynamics. Physiol Rev. 2008;88:1089–1118. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
