

Cheating is evolutionarily assimilated with cooperation in the continuous snowdrift game
- PMID: 25868940
- PMCID: PMC4441111
- DOI: 10.1016/j.biosystems.2015.04.002
Cheating is evolutionarily assimilated with cooperation in the continuous snowdrift game
Abstract
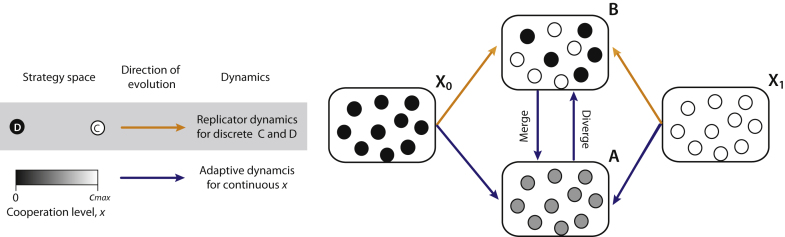
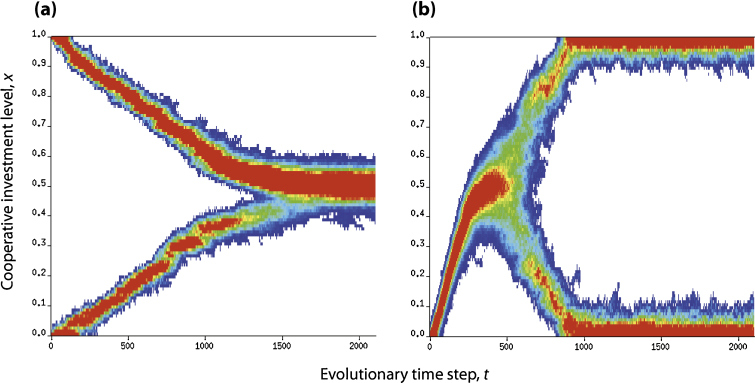
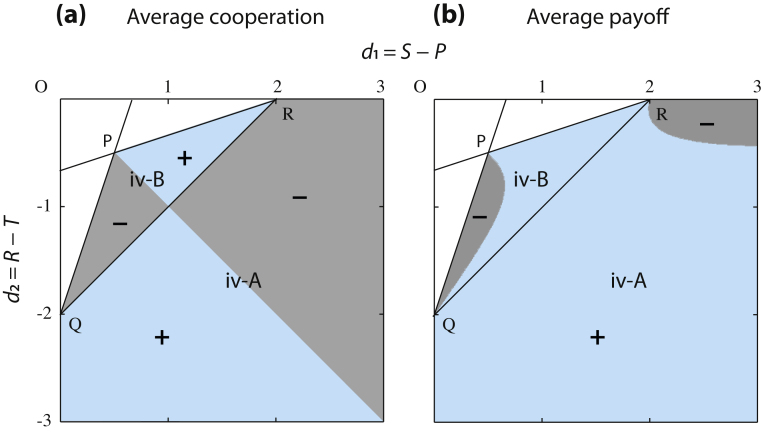
It is well known that in contrast to the Prisoner's Dilemma, the snowdrift game can lead to a stable coexistence of cooperators and cheaters. Recent theoretical evidence on the snowdrift game suggests that gradual evolution for individuals choosing to contribute in continuous degrees can result in the social diversification to a 100% contribution and 0% contribution through so-called evolutionary branching. Until now, however, game-theoretical studies have shed little light on the evolutionary dynamics and consequences of the loss of diversity in strategy. Here, we analyze continuous snowdrift games with quadratic payoff functions in dimorphic populations. Subsequently, conditions are clarified under which gradual evolution can lead a population consisting of those with 100% contribution and those with 0% contribution to merge into one species with an intermediate contribution level. The key finding is that the continuous snowdrift game is more likely to lead to assimilation of different cooperation levels rather than maintenance of diversity. Importantly, this implies that allowing the gradual evolution of cooperative behavior can facilitate social inequity aversion in joint ventures that otherwise could cause conflicts that are based on commonly accepted notions of fairness.
Keywords: Adaptive dynamics; Evolution of cooperation; Evolutionary branching; Replicator dynamics; Snowdrift game; Speciation in reverse.
Copyright © 2015 The Authors. Published by Elsevier Ireland Ltd.. All rights reserved.
Figures


References
-
- Axelrod R., Hamilton W.D. The evolution of cooperation. Science. 1981;211:1390–1396. - PubMed
-
- Axelrod R. The dissemination of culture a model with local convergence and global polarization. J. Confl. Resolut. 1997;41:203–226.
-
- Bishop D.T., Cannings C. A generalized war of attrition. J. Theor. Biol. 2015;70:85–124. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
