

The role of CD1d-restricted NKT cells in the clearance of Pseudomonas aeruginosa from the lung is dependent on the host genetic background
- PMID: 25870224
- PMCID: PMC4432766
- DOI: 10.1128/IAI.00015-15
The role of CD1d-restricted NKT cells in the clearance of Pseudomonas aeruginosa from the lung is dependent on the host genetic background
Abstract
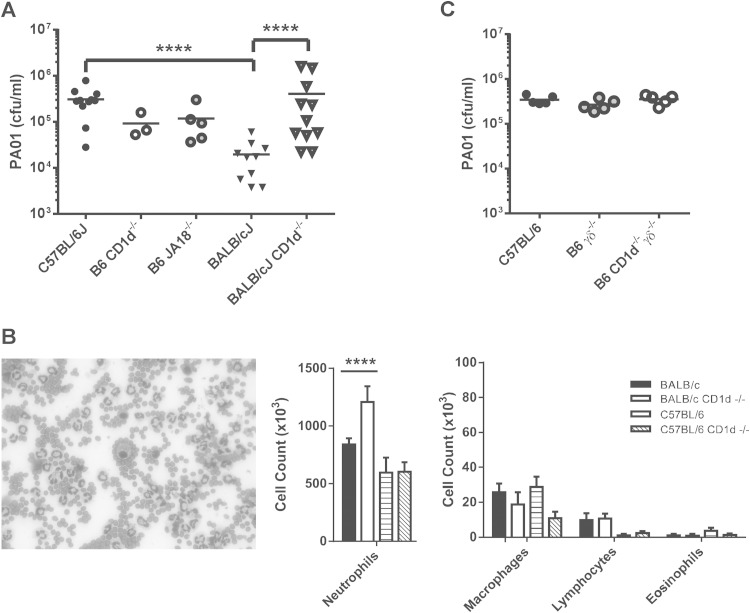
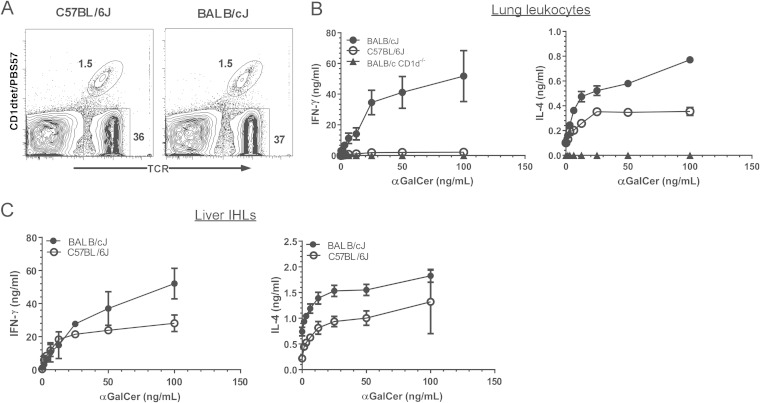
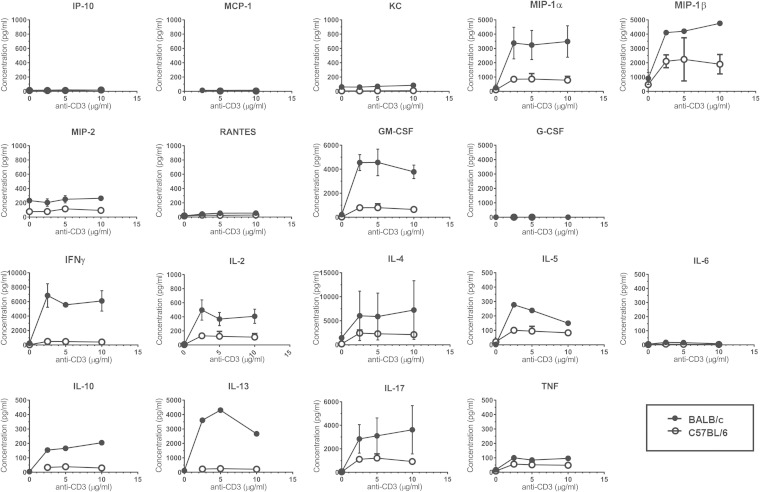
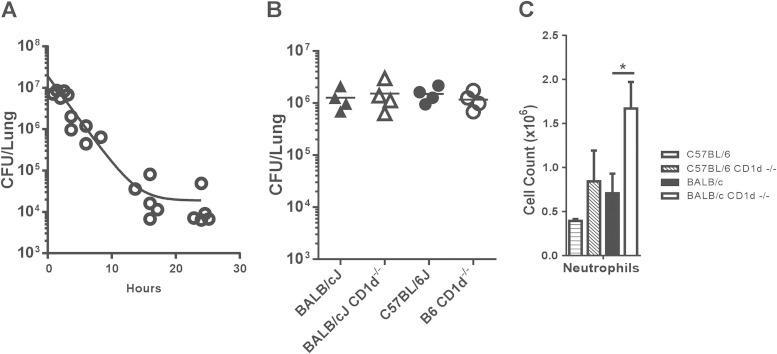
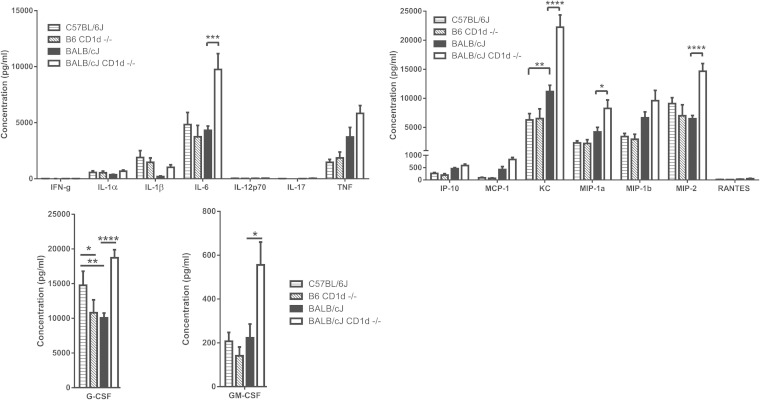
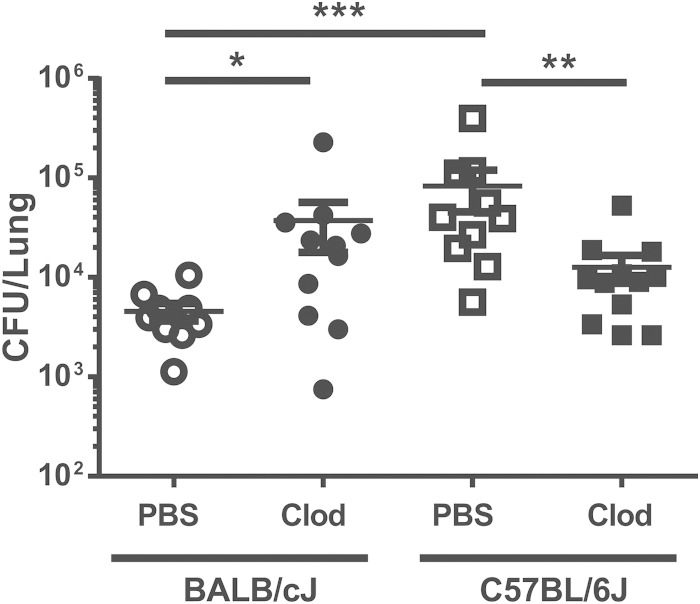
Pseudomonas aeruginosa is an important human opportunistic pathogen, accounting for a significant fraction of hospital-acquired lung infections. CD1d-restricted NKT cells comprise an unusual innate-like T cell subset that plays important roles in both bacterial and viral infections. Previous reports have differed in their conclusions regarding the role of NKT cells in clearance of P. aeruginosa from the lung. Since there is significant strain-dependent variation in NKT cell number and function among different inbred strains of mice, we investigated whether the role of NKT cells was dependent on the host genetic background. We found that NKT cells did indeed play a critical role in the clearance of P. aeruginosa from the lungs of BALB/c mice but that they played no discernible role in clearance from the lungs of C57BL/6 mice. We found that the strain-dependent role of NKT cells was associated with significant strain-dependent differences in cytokine production by lung NKT cells and that impaired clearance of P. aeruginosa in BALB/c CD1d(-/-) mice was associated with an increase in neutrophil influx to the lung and increased levels of proinflammatory cytokines and chemokines after infection. Finally, we found that the role of alveolar macrophages was also dependent on the genetic background. These data provide further support for a model in which the unusually high level of variability in NKT cell number and function among different genetic backgrounds may be an important contributor to infectious-disease susceptibility and pathology.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
