

Inhibition of TLR2 signaling by small molecule inhibitors targeting a pocket within the TLR2 TIR domain
- PMID: 25870276
- PMCID: PMC4418912
- DOI: 10.1073/pnas.1422576112
Inhibition of TLR2 signaling by small molecule inhibitors targeting a pocket within the TLR2 TIR domain
Abstract
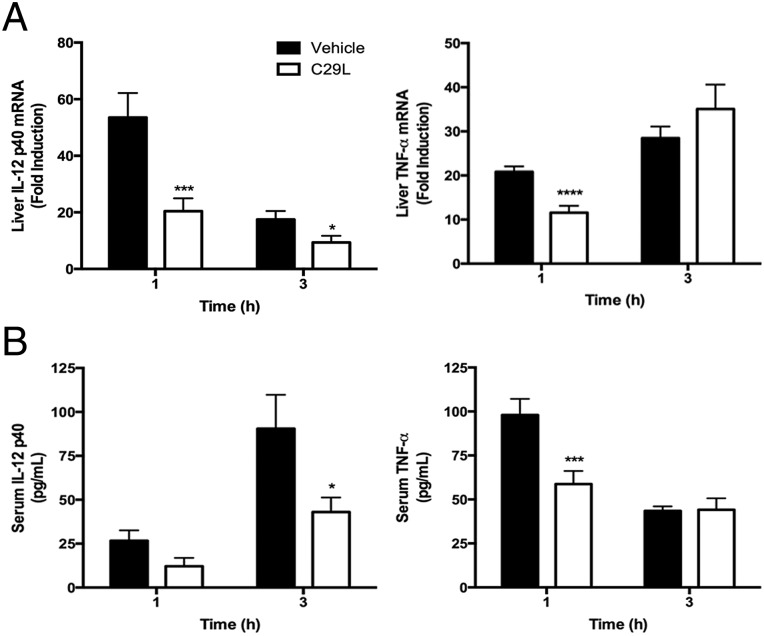
Toll-like receptor (TLR) signaling is initiated by dimerization of intracellular Toll/IL-1 receptor resistance (TIR) domains. For all TLRs except TLR3, recruitment of the adapter, myeloid differentiation primary response gene 88 (MyD88), to TLR TIR domains results in downstream signaling culminating in proinflammatory cytokine production. Therefore, blocking TLR TIR dimerization may ameliorate TLR2-mediated hyperinflammatory states. The BB loop within the TLR TIR domain is critical for mediating certain protein-protein interactions. Examination of the human TLR2 TIR domain crystal structure revealed a pocket adjacent to the highly conserved P681 and G682 BB loop residues. Using computer-aided drug design (CADD), we sought to identify a small molecule inhibitor(s) that would fit within this pocket and potentially disrupt TLR2 signaling. In silico screening identified 149 compounds and 20 US Food and Drug Administration-approved drugs based on their predicted ability to bind in the BB loop pocket. These compounds were screened in HEK293T-TLR2 transfectants for the ability to inhibit TLR2-mediated IL-8 mRNA. C16H15NO4 (C29) was identified as a potential TLR2 inhibitor. C29, and its derivative, ortho-vanillin (o-vanillin), inhibited TLR2/1 and TLR2/6 signaling induced by synthetic and bacterial TLR2 agonists in human HEK-TLR2 and THP-1 cells, but only TLR2/1 signaling in murine macrophages. C29 failed to inhibit signaling induced by other TLR agonists and TNF-α. Mutagenesis of BB loop pocket residues revealed an indispensable role for TLR2/1, but not TLR2/6, signaling, suggesting divergent roles. Mice treated with o-vanillin exhibited reduced TLR2-induced inflammation. Our data provide proof of principle that targeting the BB loop pocket is an effective approach for identification of TLR2 signaling inhibitors.
Keywords: BB loop; CADD; TLR2 pocket; small molecule inhibitor.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Medzhitov R. Toll-like receptors and innate immunity. Nat Rev Immunol. 2001;1(2):135–145. - PubMed
-
- Takeuchi O, et al. Discrimination of bacterial lipoproteins by Toll-like receptor 6. Int Immunol. 2001;13(7):933–940. - PubMed
-
- Takeuchi O, et al. Cutting edge: Role of Toll-like receptor 1 in mediating immune response to microbial lipoproteins. J Immunol. 2002;169(1):10–14. - PubMed
-
- Cole LE, et al. Immunologic consequences of Francisella tularensis live vaccine strain infection: Role of the innate immune response in infection and immunity. J Immunol. 2006;176(11):6888–6899. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
