

Host attachment and fluid shear are integrated into a mechanical signal regulating virulence in Escherichia coli O157:H7
- PMID: 25870295
- PMCID: PMC4418854
- DOI: 10.1073/pnas.1422986112
Host attachment and fluid shear are integrated into a mechanical signal regulating virulence in Escherichia coli O157:H7
Abstract
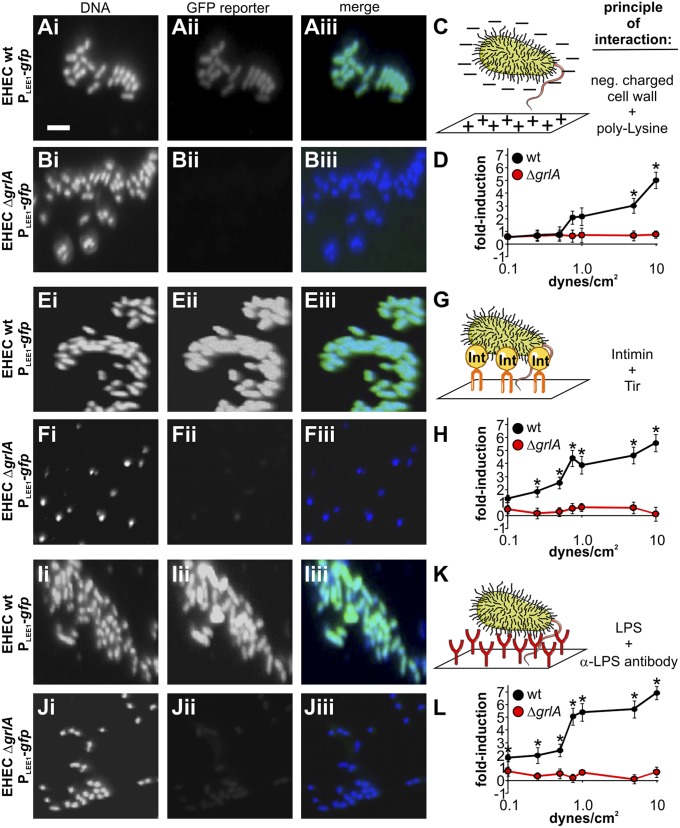
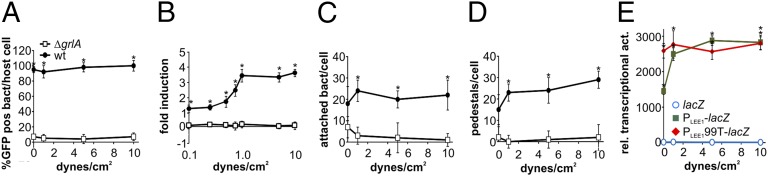
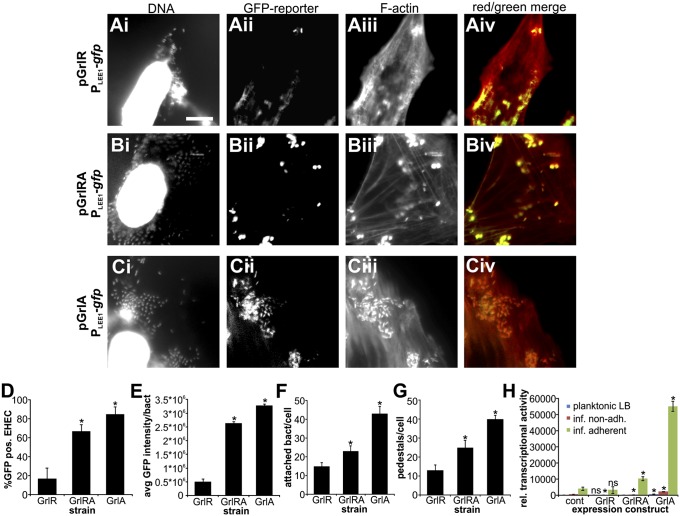
Enterohemorrhagic Escherichia coli (EHEC) is a foodborne pathogen causing hemorrhagic colitis and hemolytic uremic syndrome. EHEC colonizes the intestinal tract through a range of virulence factors encoded by the locus of enterocyte effacement (LEE), as well as Shiga toxin. Although the factors involved in colonization and disease are well characterized, how EHEC regulates its expression in response to a host encounter is not well understood. Here, we report that EHEC perceives attachment to host cells as a mechanical cue that leads to expression of LEE-encoded virulence genes. This signal is transduced via the LEE-encoded global regulator of LEE-encoded regulator (Ler) and global regulator of Ler and is further enhanced by levels of shear force similar to peristaltic forces in the intestinal tract. Our data suggest that, in addition to a range of chemical environmental signals, EHEC is capable of sensing and responding to mechanical cues to adapt to its host's physiology.
Keywords: attaching/effacing pathogens; enterohemorrhagic Escherichia coli; gastrointestinal infection; locus of enterocyte effacement; mechanosensing.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
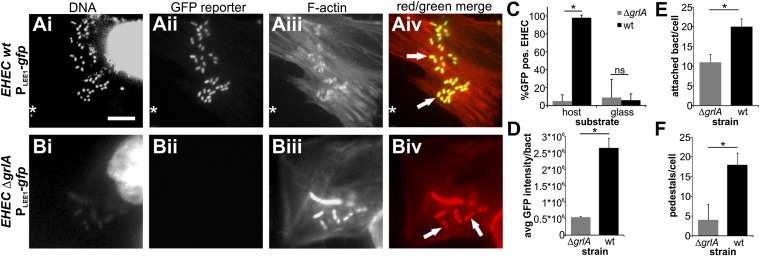
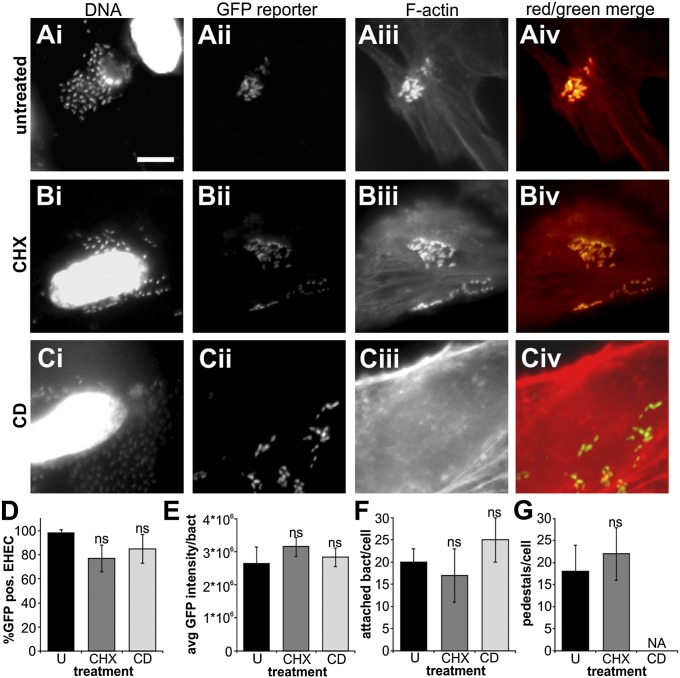
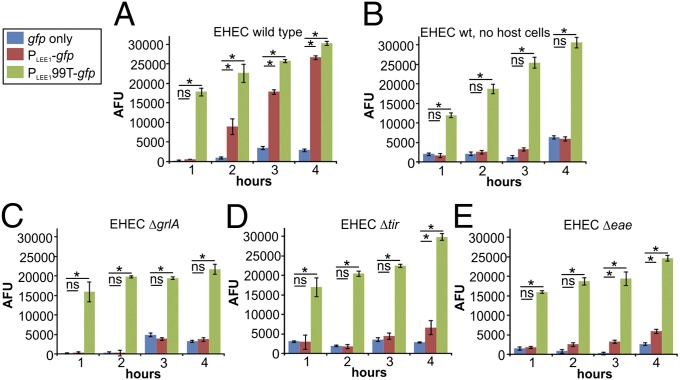
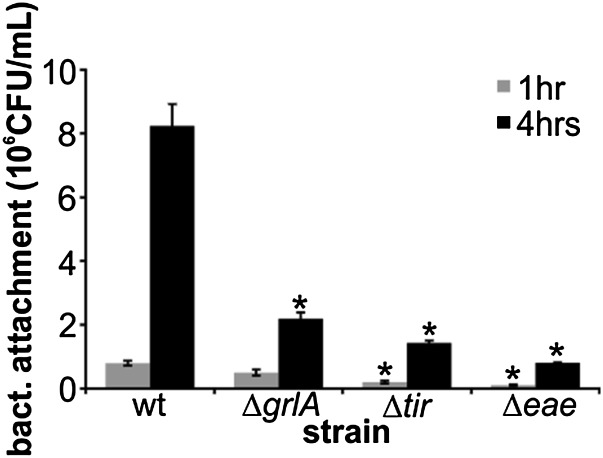
References
-
- McDaniel TK, Kaper JB. A cloned pathogenicity island from enteropathogenic Escherichia coli confers the attaching and effacing phenotype on E. coli K-12. Mol Microbiol. 1997;23(2):399–407. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
