

A general framework for thermodynamically consistent parameterization and efficient sampling of enzymatic reactions
- PMID: 25874556
- PMCID: PMC4397067
- DOI: 10.1371/journal.pcbi.1004195
A general framework for thermodynamically consistent parameterization and efficient sampling of enzymatic reactions
Abstract
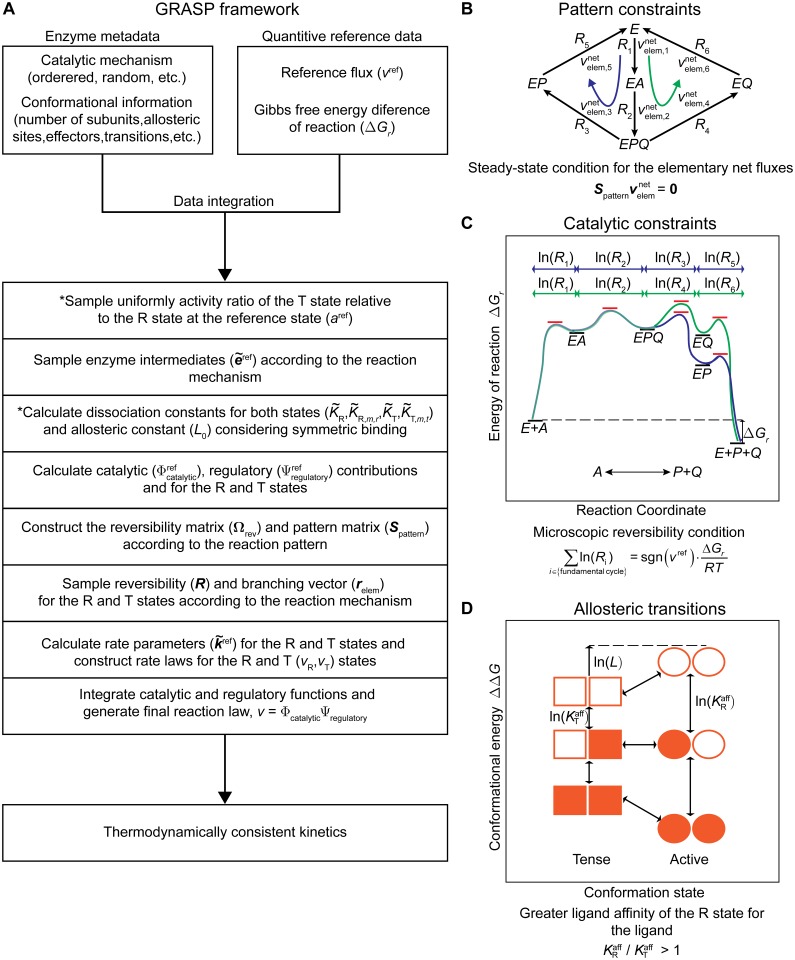
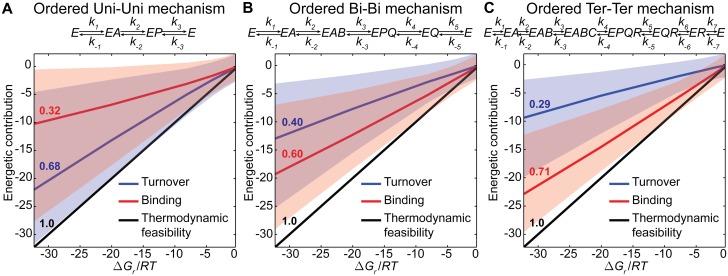
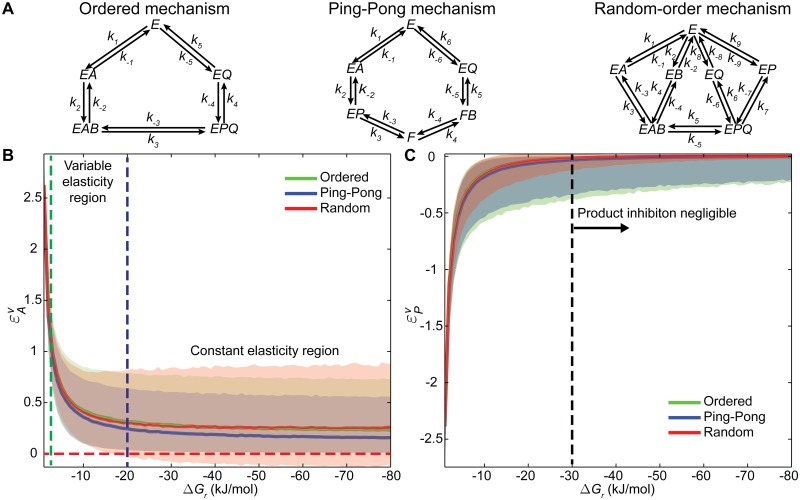
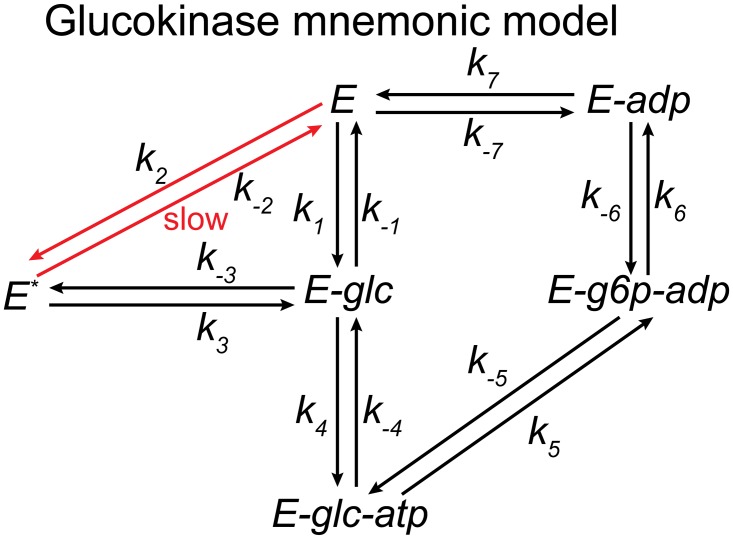
Kinetic models provide the means to understand and predict the dynamic behaviour of enzymes upon different perturbations. Despite their obvious advantages, classical parameterizations require large amounts of data to fit their parameters. Particularly, enzymes displaying complex reaction and regulatory (allosteric) mechanisms require a great number of parameters and are therefore often represented by approximate formulae, thereby facilitating the fitting but ignoring many real kinetic behaviours. Here, we show that full exploration of the plausible kinetic space for any enzyme can be achieved using sampling strategies provided a thermodynamically feasible parameterization is used. To this end, we developed a General Reaction Assembly and Sampling Platform (GRASP) capable of consistently parameterizing and sampling accurate kinetic models using minimal reference data. The former integrates the generalized MWC model and the elementary reaction formalism. By formulating the appropriate thermodynamic constraints, our framework enables parameterization of any oligomeric enzyme kinetics without sacrificing complexity or using simplifying assumptions. This thermodynamically safe parameterization relies on the definition of a reference state upon which feasible parameter sets can be efficiently sampled. Uniform sampling of the kinetics space enabled dissecting enzyme catalysis and revealing the impact of thermodynamics on reaction kinetics. Our analysis distinguished three reaction elasticity regions for common biochemical reactions: a steep linear region (0> ΔGr >-2 kJ/mol), a transition region (-2> ΔGr >-20 kJ/mol) and a constant elasticity region (ΔGr <-20 kJ/mol). We also applied this framework to model more complex kinetic behaviours such as the monomeric cooperativity of the mammalian glucokinase and the ultrasensitive response of the phosphoenolpyruvate carboxylase of Escherichia coli. In both cases, our approach described appropriately not only the kinetic behaviour of these enzymes, but it also provided insights about the particular features underpinning the observed kinetics. Overall, this framework will enable systematic parameterization and sampling of enzymatic reactions.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
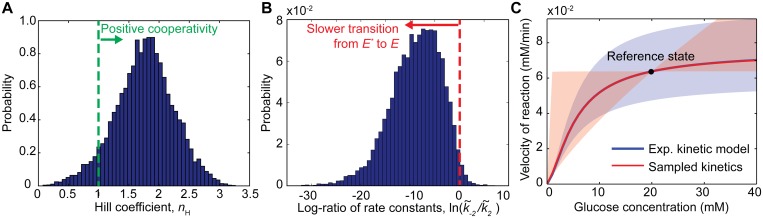
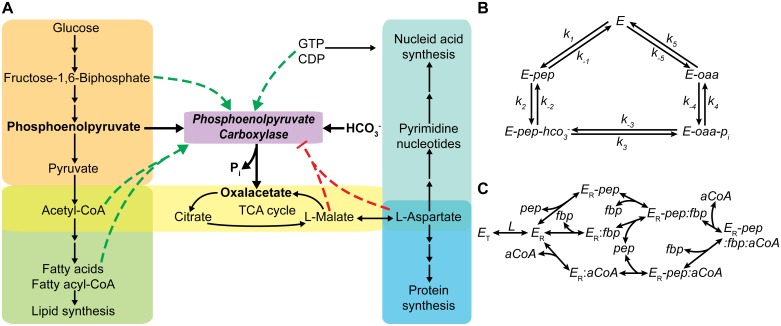
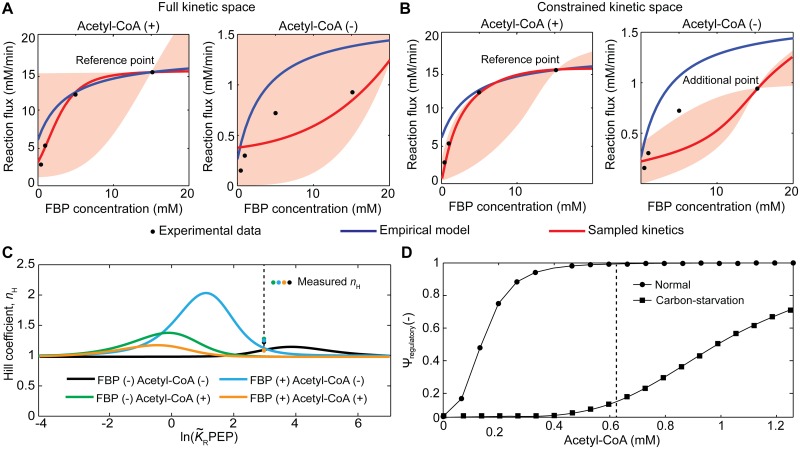
References
-
- Michaelis L, Menten ML (1913) Die Kinetik der Invertinwirkung. Biochemische Zeitschrift 49: 333–369.
-
- Cornish-Bowden A (2012) Fundamentals of Enzyme Kinetics: Wiley-Blackwell.
-
- Alberty R (2006) Relations between biochemical thermodynamics and biochemical kinetics. Biophys Chem 124: 11–17. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
