

Relating structure and function of inner hair cell ribbon synapses
- PMID: 25874597
- PMCID: PMC4487357
- DOI: 10.1007/s00441-014-2102-7
Relating structure and function of inner hair cell ribbon synapses
Abstract
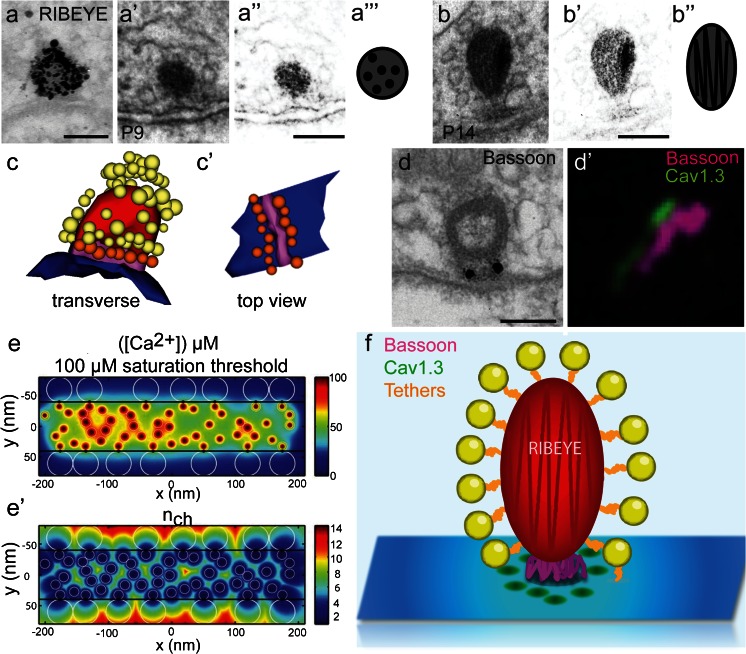
In the mammalian cochlea, sound is encoded at synapses between inner hair cells (IHCs) and type I spiral ganglion neurons (SGNs). Each SGN receives input from a single IHC ribbon-type active zone (AZ) and yet SGNs indefatigably spike up to hundreds of Hz to encode acoustic stimuli with submillisecond precision. Accumulating evidence indicates a highly specialized molecular composition and structure of the presynapse, adapted to suit these high functional demands. However, we are only beginning to understand key features such as stimulus-secretion coupling, exocytosis mechanisms, exo-endocytosis coupling, modes of endocytosis and vesicle reformation, as well as replenishment of the readily releasable pool. Relating structure and function has become an important avenue in addressing these points and has been applied to normal and genetically manipulated hair cell synapses. Here, we review some of the exciting new insights gained from recent studies of the molecular anatomy and physiology of IHC ribbon synapses.
Figures


References
-
- Altrock WD, et al. Functional inactivation of a fraction of excitatory synapses in mice deficient for the active zone protein bassoon. Neuron. 2003;37:787–800. - PubMed
-
- Betz A, Ashery U, Rickmann M, Augustin I, Neher E, Südhof TC, Rettig J, Brose N. Munc13-1 is a presynaptic phorbol ester receptor that enhances neurotransmitter release. Neuron. 1998;21:123–136. - PubMed
-
- Betz A, Thakur P, Junge HJ, Ashery U, Rhee J-S, Scheuss V, Rosenmund C, Rettig J, Brose N. Functional interaction of the active zone proteins Munc13-1 and RIM1 in synaptic vesicle priming. Neuron. 2001;30:183–196. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
