

A critical period for postnatal adaptive plasticity in a model of motor axon miswiring
- PMID: 25874621
- PMCID: PMC4395312
- DOI: 10.1371/journal.pone.0123643
A critical period for postnatal adaptive plasticity in a model of motor axon miswiring
Abstract
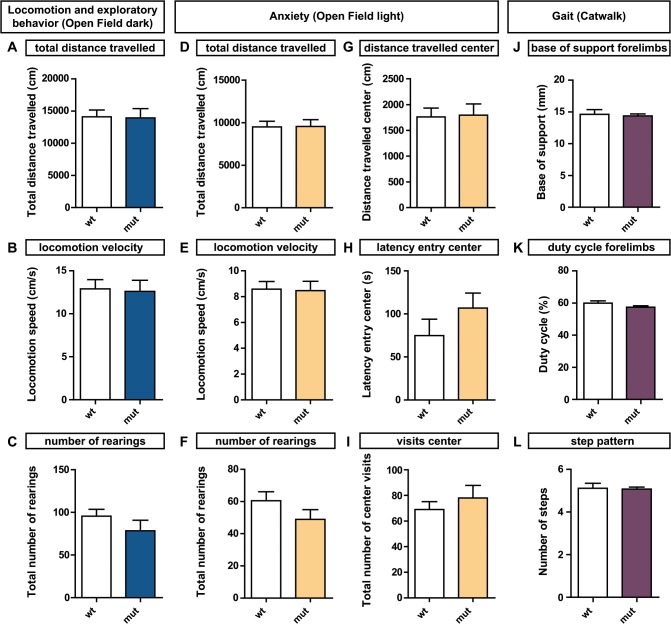
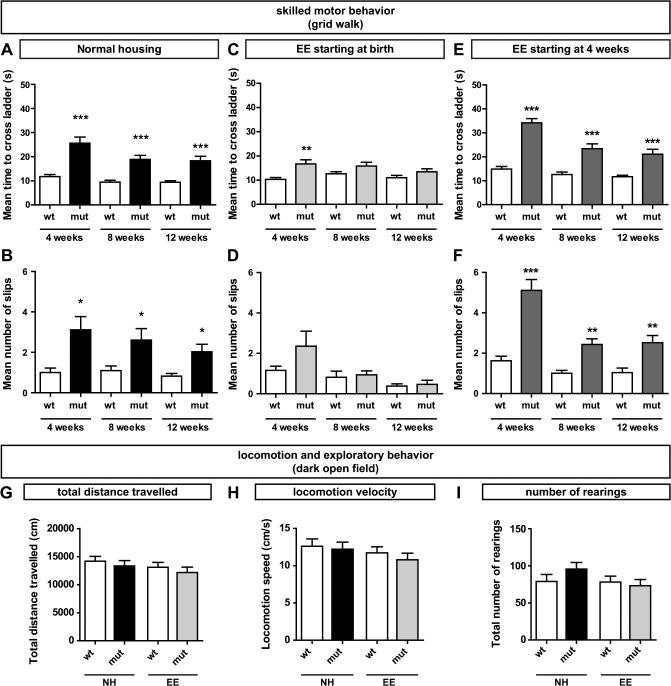
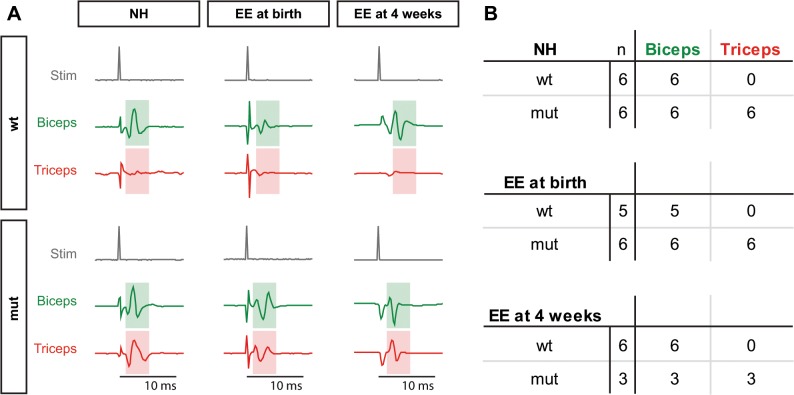
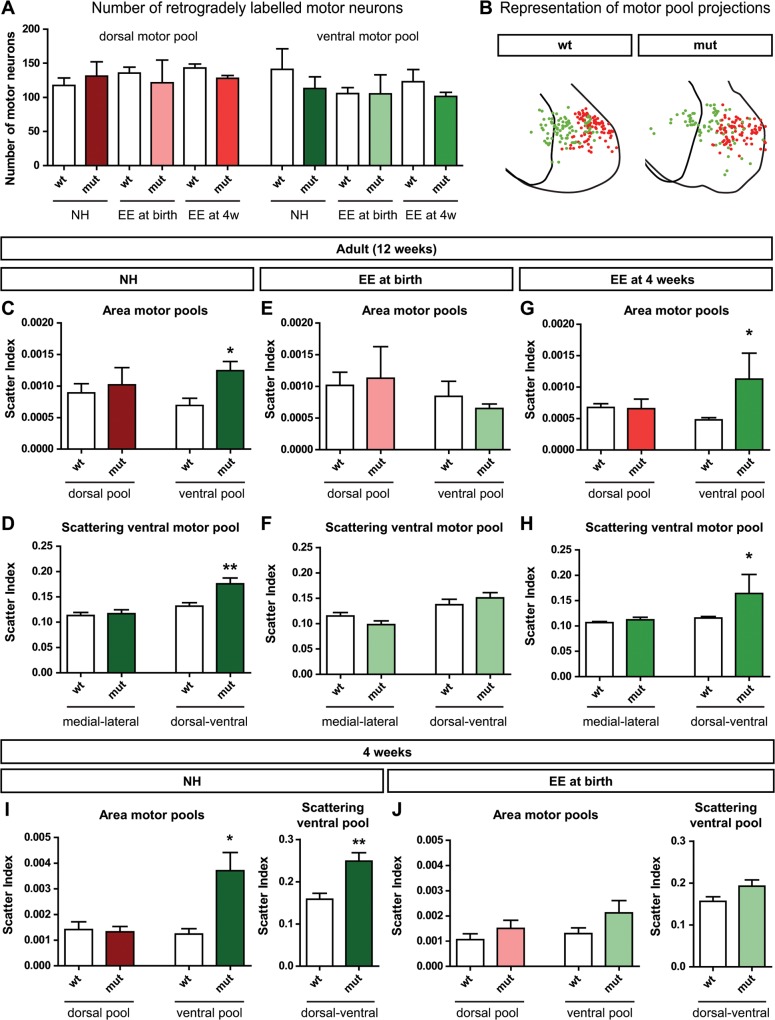
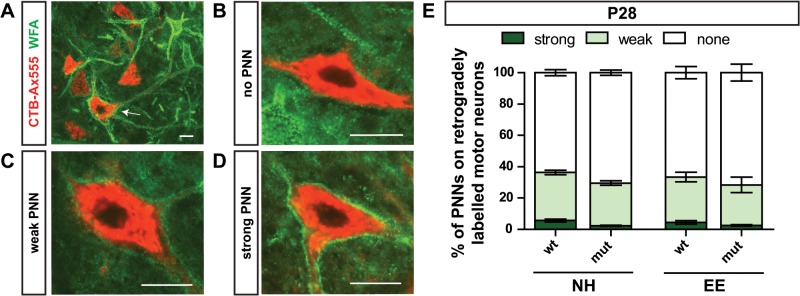
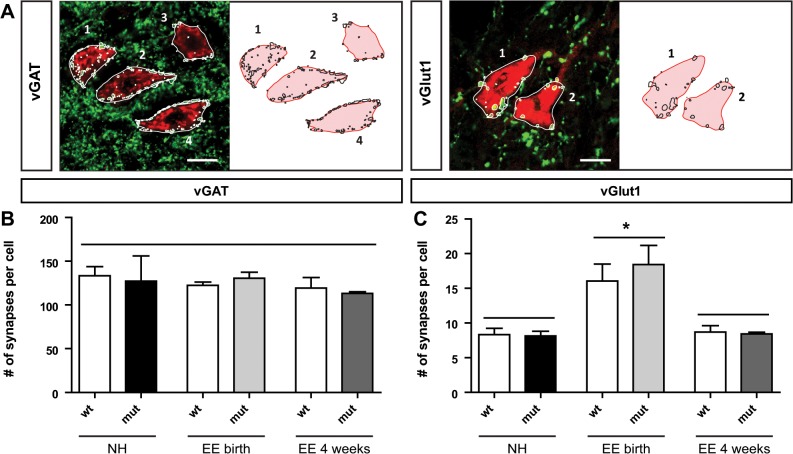
The correct wiring of neuronal circuits is of crucial importance for precise neuromuscular functionality. Therefore, guidance cues provide tight spatiotemporal control of axon growth and guidance. Mice lacking the guidance cue Semaphorin 3F (Sema3F) display very specific axon wiring deficits of motor neurons in the medial aspect of the lateral motor column (LMCm). While these deficits have been investigated extensively during embryonic development, it remained unclear how Sema3F mutant mice cope with these errors postnatally. We therefore investigated whether these animals provide a suitable model for the exploration of adaptive plasticity in a system of miswired neuronal circuitry. We show that the embryonically developed wiring deficits in Sema3F mutants persist until adulthood. As a consequence, these mutants display impairments in motor coordination that improve during normal postnatal development, but never reach wildtype levels. These improvements in motor coordination were boosted to wildtype levels by housing the animals in an enriched environment starting at birth. In contrast, a delayed start of enriched environment housing, at 4 weeks after birth, did not similarly affect motor performance of Sema3F mutants. These results, which are corroborated by neuroanatomical analyses, suggest a critical period for adaptive plasticity in neuromuscular circuitry. Interestingly, the formation of perineuronal nets, which are known to close the critical period for plastic changes in other systems, was not altered between the different housing groups. However, we found significant changes in the number of excitatory synapses on limb innervating motor neurons. Thus, we propose that during the early postnatal phase, when perineuronal nets have not yet been formed around spinal motor neurons, housing in enriched environment conditions induces adaptive plasticity in the motor system by the formation of additional synaptic contacts, in order to compensate for coordination deficits.
Conflict of interest statement
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
