

Fourth-generation epac-based FRET sensors for cAMP feature exceptional brightness, photostability and dynamic range: characterization of dedicated sensors for FLIM, for ratiometry and with high affinity
- PMID: 25875503
- PMCID: PMC4397040
- DOI: 10.1371/journal.pone.0122513
Fourth-generation epac-based FRET sensors for cAMP feature exceptional brightness, photostability and dynamic range: characterization of dedicated sensors for FLIM, for ratiometry and with high affinity
Abstract
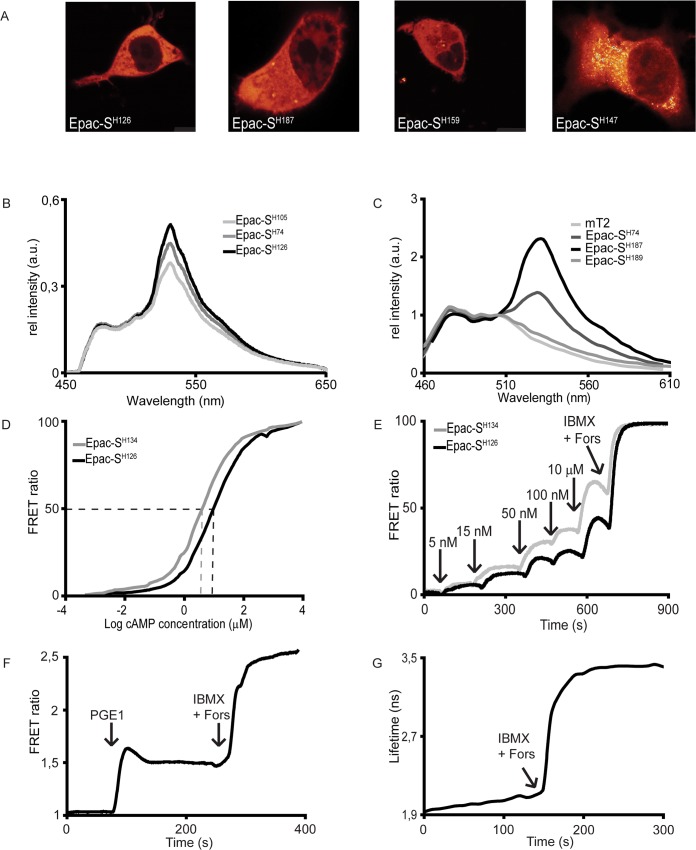
Epac-based FRET sensors have been widely used for the detection of cAMP concentrations in living cells. Originally developed by us as well as others, we have since then reported several important optimizations that make these sensors favourite among many cell biologists. We here report cloning and characterization of our fourth generation of cAMP sensors, which feature outstanding photostability, dynamic range and signal-to-noise ratio. The design is based on mTurquoise2, currently the brightest and most bleaching-resistant donor, and a new acceptor cassette that consists of a tandem of two cp173Venus fluorophores. We also report variants with a single point mutation, Q270E, in the Epac moiety, which decreases the dissociation constant of cAMP from 9.5 to 4 μM, and thus increases the affinity ~ 2.5-fold. Finally, we also prepared and characterized dedicated variants with non-emitting (dark) acceptors for single-wavelength FLIM acquisition that display an exceptional near-doubling of fluorescence lifetime upon saturation of cAMP levels. We believe this generation of cAMP outperforms all other sensors and therefore recommend these sensors for all future studies.
Conflict of interest statement
Figures

Similar articles
-
Cytosolic-enhanced dark Epac-based FRET sensors allow for intracellular cAMP detection in live cells via FLIM.FEBS Lett. 2025 Apr;599(7):1075-1085. doi: 10.1002/1873-3468.15093. Epub 2024 Dec 31. FEBS Lett. 2025. PMID: 39737677 Free PMC article.
-
Dynamic monitoring of Gi/o-protein-mediated decreases of intracellular cAMP by FRET-based Epac sensors.Pflugers Arch. 2017 Jun;469(5-6):725-737. doi: 10.1007/s00424-017-1975-1. Epub 2017 Apr 6. Pflugers Arch. 2017. PMID: 28386636 Free PMC article.
-
A mTurquoise-based cAMP sensor for both FLIM and ratiometric read-out has improved dynamic range.PLoS One. 2011 Apr 29;6(4):e19170. doi: 10.1371/journal.pone.0019170. PLoS One. 2011. PMID: 21559477 Free PMC article.
-
Interrogating cyclic AMP signaling using optical approaches.Cell Calcium. 2017 Jun;64:47-56. doi: 10.1016/j.ceca.2017.02.010. Epub 2017 Mar 1. Cell Calcium. 2017. PMID: 28274483 Free PMC article. Review.
-
Quantitative Live Cell FLIM Imaging in Three Dimensions.Adv Exp Med Biol. 2017;1035:31-48. doi: 10.1007/978-3-319-67358-5_3. Adv Exp Med Biol. 2017. PMID: 29080129 Review.
Cited by
-
Phosphodiesterase 1C integrates store-operated calcium entry and cAMP signaling in leading-edge protrusions of migrating human arterial myocytes.J Biol Chem. 2021 Jan-Jun;296:100606. doi: 10.1016/j.jbc.2021.100606. Epub 2021 Mar 28. J Biol Chem. 2021. PMID: 33789162 Free PMC article.
-
Estimating the magnitude of near-membrane PDE4 activity in living cells.Am J Physiol Cell Physiol. 2015 Sep 15;309(6):C415-24. doi: 10.1152/ajpcell.00090.2015. Epub 2015 Jul 22. Am J Physiol Cell Physiol. 2015. PMID: 26201952 Free PMC article.
-
cAMP Bursts Control T Cell Directionality by Actomyosin Cytoskeleton Remodeling.Front Cell Dev Biol. 2021 May 20;9:633099. doi: 10.3389/fcell.2021.633099. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34095108 Free PMC article.
-
Myocardial TRPC6-mediated Zn2+ influx induces beneficial positive inotropy through β-adrenoceptors.Nat Commun. 2022 Oct 26;13(1):6374. doi: 10.1038/s41467-022-34194-9. Nat Commun. 2022. PMID: 36289215 Free PMC article.
-
Human nonvisual opsin 3 regulates pigmentation of epidermal melanocytes through functional interaction with melanocortin 1 receptor.Proc Natl Acad Sci U S A. 2019 Jun 4;116(23):11508-11517. doi: 10.1073/pnas.1902825116. Epub 2019 May 16. Proc Natl Acad Sci U S A. 2019. PMID: 31097585 Free PMC article.
References
-
- Nikolaev VO, Bunemann M, Hein L, Hannawacker A, Lohse MJ. Novel single chain cAMP sensors for receptor-induced signal propagation. J Biol Chem 2004;279: 37215–37218. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
