

A mathematical model of collective cell migration in a three-dimensional, heterogeneous environment
- PMID: 25875645
- PMCID: PMC4395426
- DOI: 10.1371/journal.pone.0122799
A mathematical model of collective cell migration in a three-dimensional, heterogeneous environment
Abstract
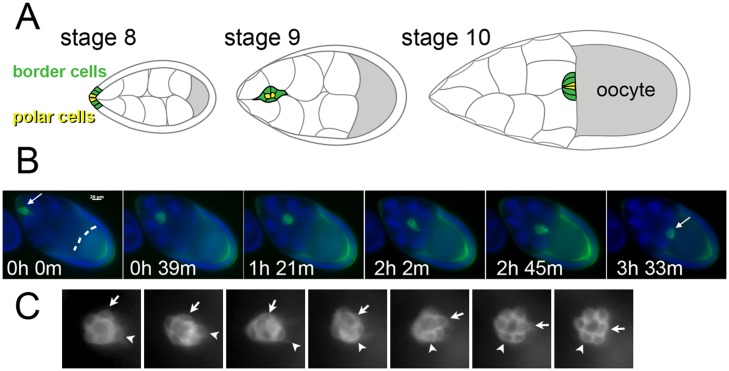
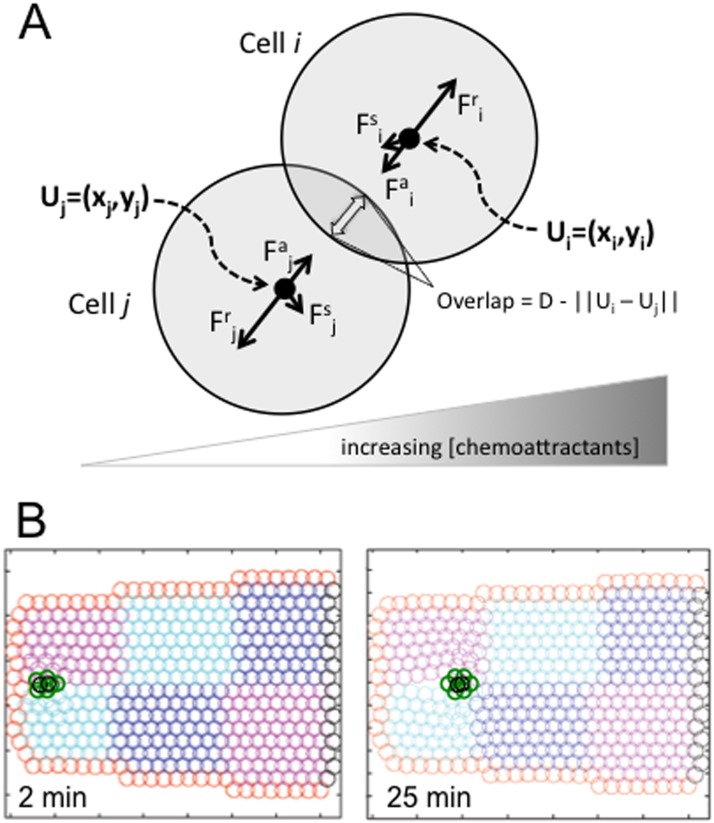
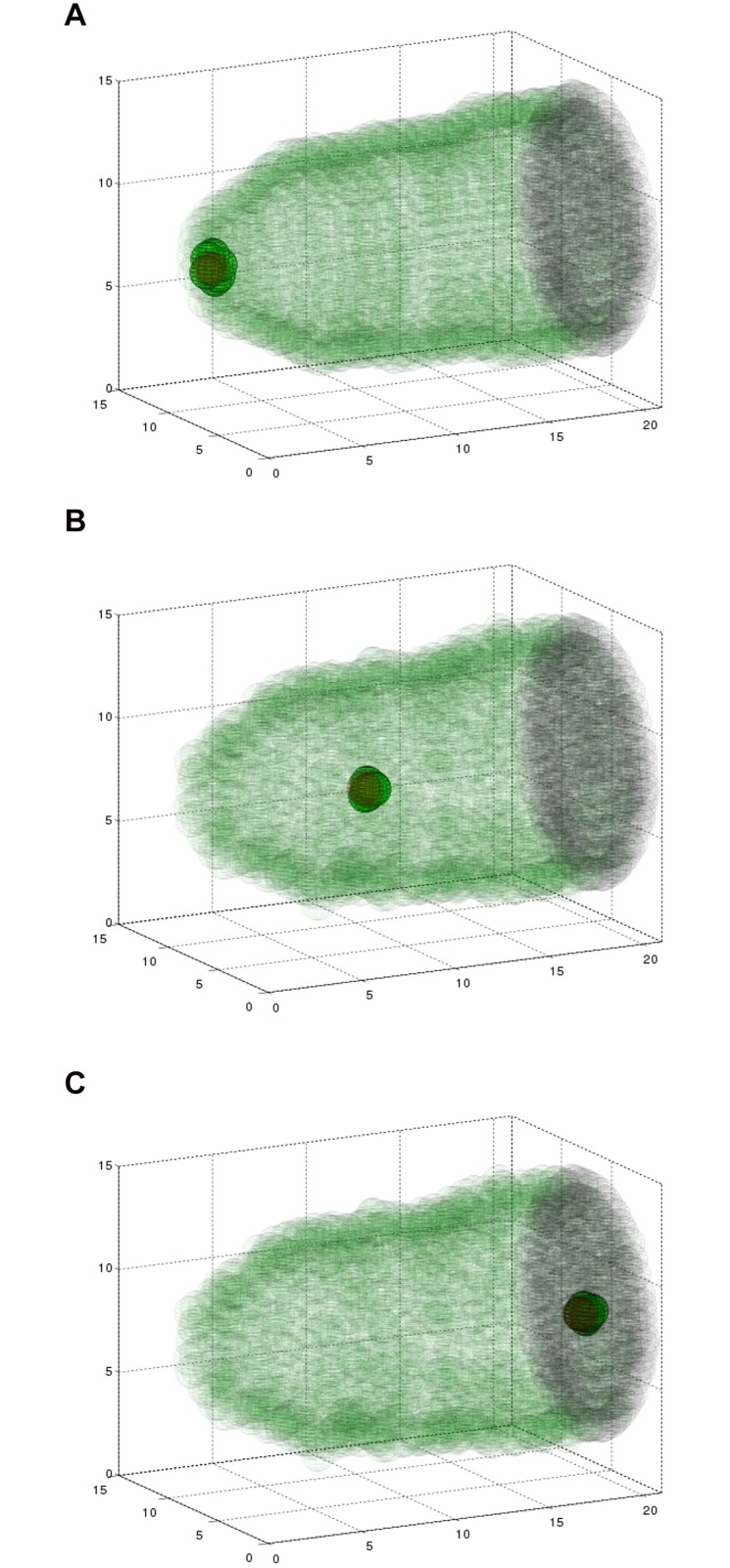
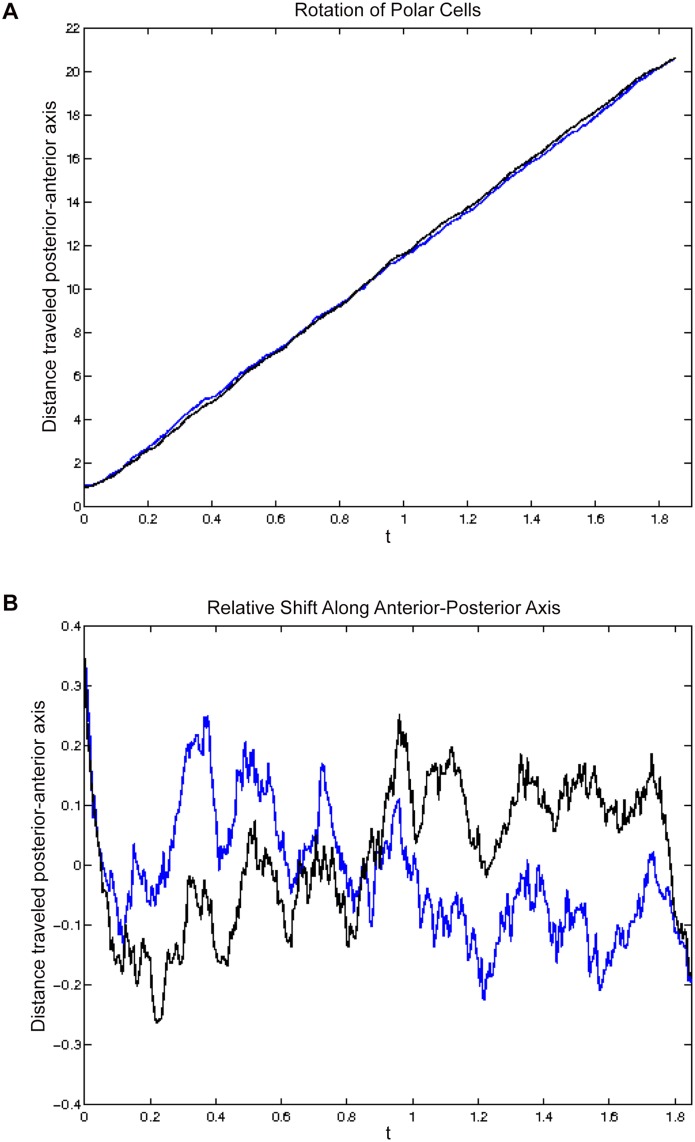
Cell migration is essential in animal development, homeostasis, and disease progression, but many questions remain unanswered about how this process is controlled. While many kinds of individual cell movements have been characterized, less effort has been directed towards understanding how clusters of cells migrate collectively through heterogeneous, cellular environments. To explore this, we have focused on the migration of the border cells during Drosophila egg development. In this case, a cluster of different cell types coalesce and traverse as a group between large cells, called nurse cells, in the center of the egg chamber. We have developed a new model for this collective cell migration based on the forces of adhesion, repulsion, migration and stochastic fluctuation to generate the movement of discrete cells. We implement the model using Identical Math Cells, or IMCs. IMCs can each represent one biological cell of the system, or can be aggregated using increased adhesion forces to model the dynamics of larger biological cells. The domain of interest is filled with IMCs, each assigned specific biophysical properties to mimic a diversity of cell types. Using this system, we have successfully simulated the migration of the border cell cluster through an environment filled with larger cells, which represent nurse cells. Interestingly, our simulations suggest that the forces utilized in this model are sufficient to produce behaviors of the cluster that are observed in vivo, such as rotation. Our framework was developed to capture a heterogeneous cell population, and our implementation strategy allows for diverse, but precise, initial position specification over a three- dimensional domain. Therefore, we believe that this model will be useful for not only examining aspects of Drosophila oogenesis, but also for modeling other two or three-dimensional systems that have multiple cell types and where investigating the forces between cells is of interest.
Conflict of interest statement
Figures
References
-
- Starz-Gaiano M, Melani M, Wang X, Meinhardt H, Montell DJ. Feedback Inhibition of JAK/STAT Signaling by Apontic Is Required to Limit an Invasive Cell Population. Developmental Cell. 2008;14(5):726–738. Available from: http://www.sciencedirect.com/science/article/pii/S1534580708001135 . doi: http://www.sciencedirect.com/science/article/pii/S1534580708001135 10.1016/j.devcel.2008.03.005. doi: 10.1016/j.devcel.2008.03.005 - DOI - PubMed
-
- Ridley AJ, Schwartz MA, Burridge K, Firtel RA, Ginsberg MH, Borisy G, et al. Cell Migration: Integrating Signals from Front to Back. Science. 2003;302(5651):1704–1709. Available from: http://www.sciencemag.org/content/302/5651/1704.abstract . doi: http://www.sciencemag.org/content/302/5651/1704.abstract 10.1126/science.1092053. doi: 10.1126/science.1092053 - DOI - PubMed
-
- Wright N, Hoffmann W, Otto W, Rio MC, Thim L. Rolling in the clover: trefoil factor family (TFF)-domain peptides, cell migration and cancer. FEBS Letters. 1997;408(2):121–123. Available from: http://www.sciencedirect.com/science/article/pii/S0014579397004249 . doi: http://www.sciencedirect.com/science/article/pii/S0014579397004249 10.1016/S0014-5793(97)00424-9. doi: 10.1016/S0014-5793(97)00424-9 - DOI - PubMed
-
- Ridley AJ. Rho GTPases and cell migration. Journal of Cell Science. 2001;114(15):2713–2722. Available from: http://jcs.biologists.org/content/114/15/2713.abstract - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
