

STING agonist formulated cancer vaccines can cure established tumors resistant to PD-1 blockade
- PMID: 25877890
- PMCID: PMC4504692
- DOI: 10.1126/scitranslmed.aaa4306
STING agonist formulated cancer vaccines can cure established tumors resistant to PD-1 blockade
Abstract
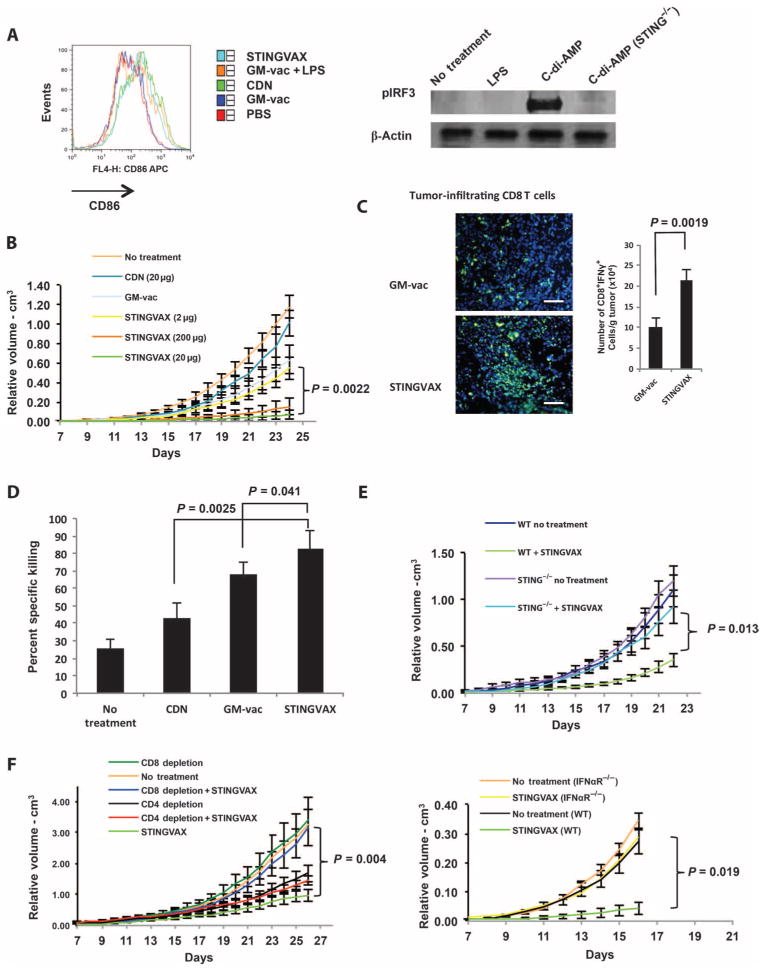
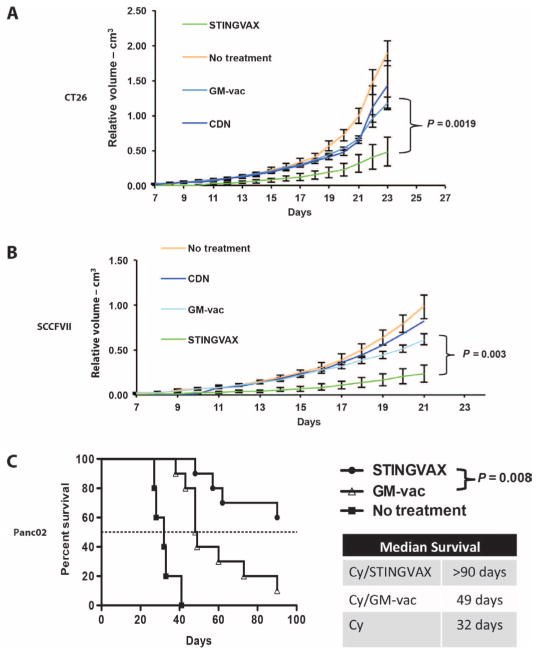
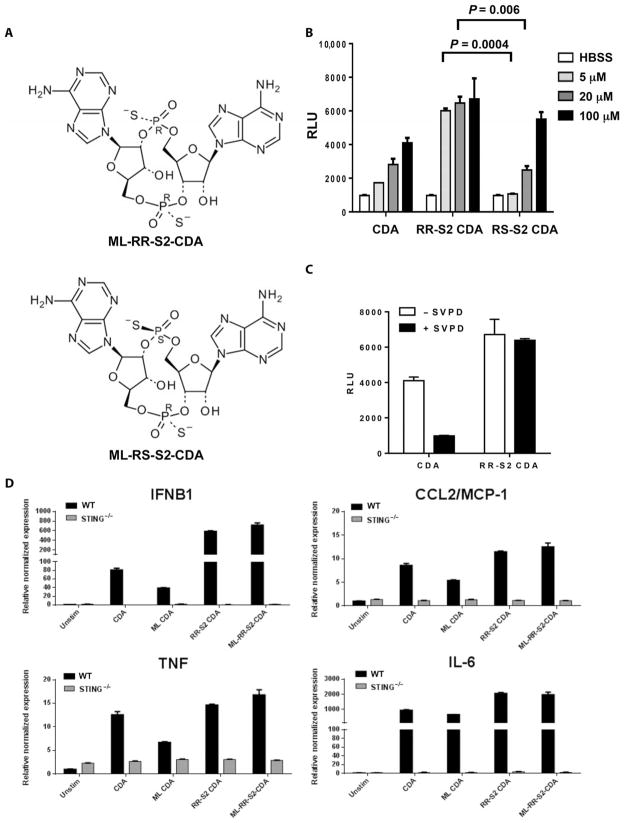
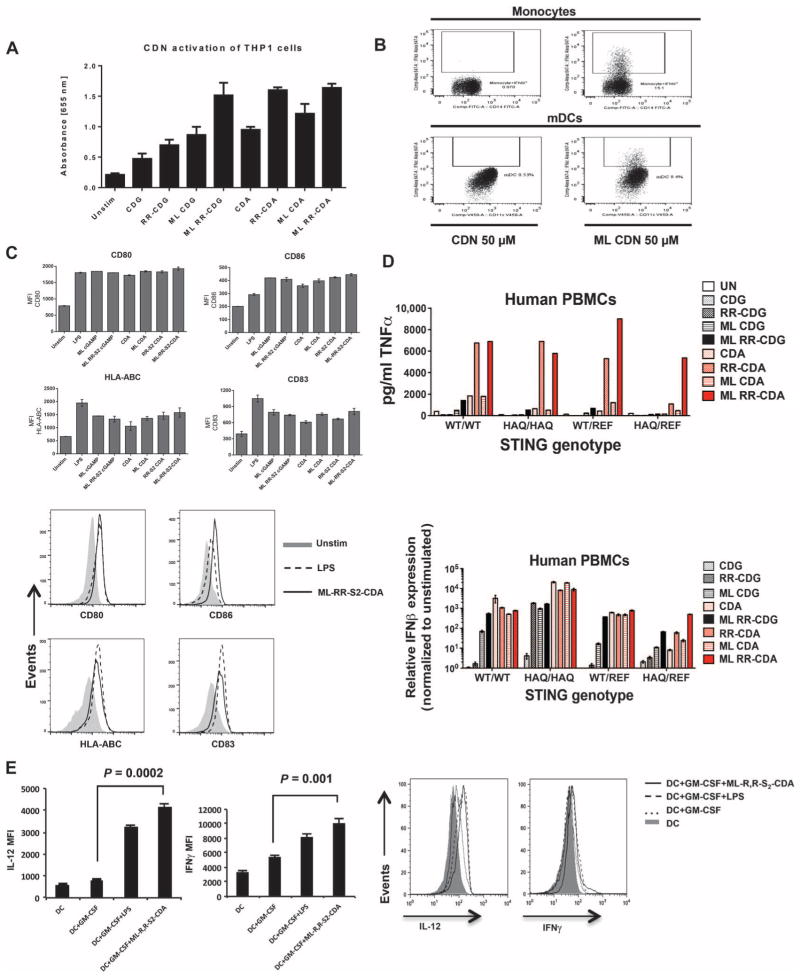
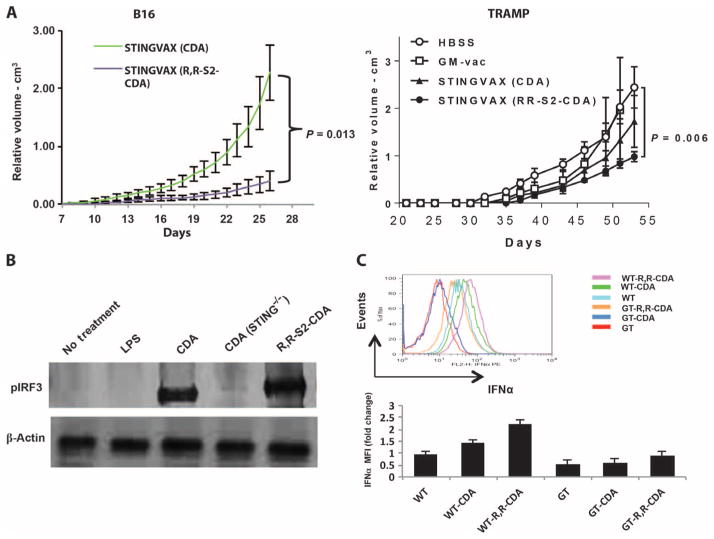
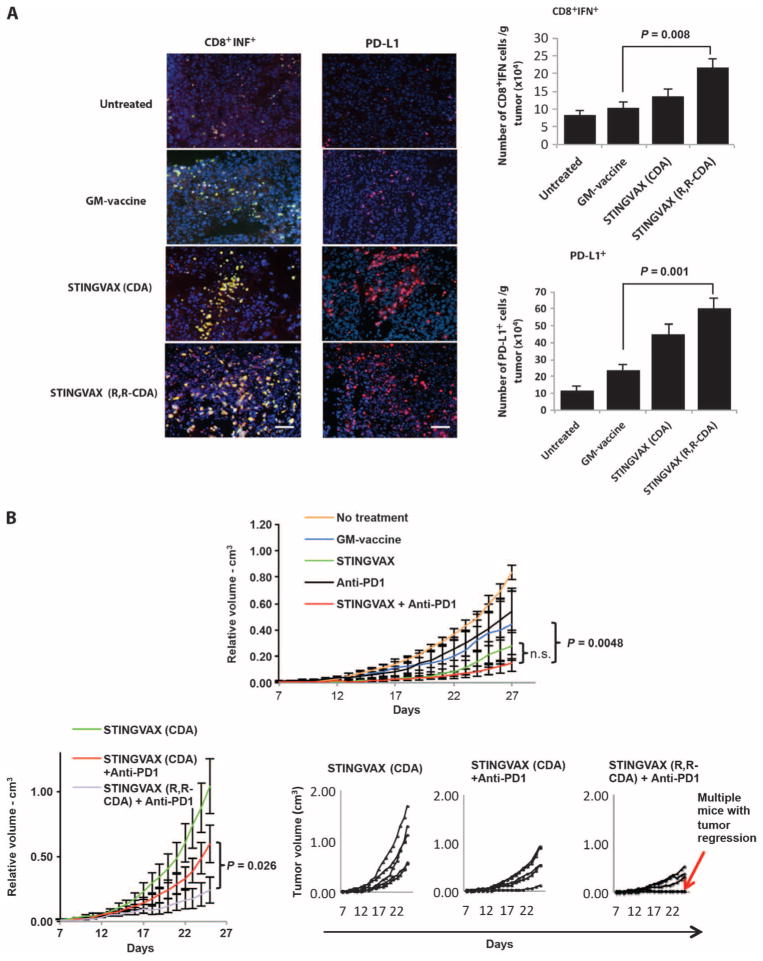
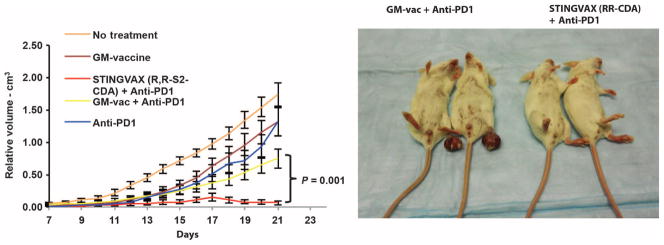
Stimulator of interferon genes (STING) is a cytosolic receptor that senses both exogenous and endogenous cytosolic cyclic dinucleotides (CDNs), activating TBK1/IRF3 (interferon regulatory factor 3), NF-κB (nuclear factor κB), and STAT6 (signal transducer and activator of transcription 6) signaling pathways to induce robust type I interferon and proinflammatory cytokine responses. CDN ligands were formulated with granulocyte-macrophage colony-stimulating factor (GM-CSF)-producing cellular cancer vaccines--termed STINGVAX--that demonstrated potent in vivo antitumor efficacy in multiple therapeutic models of established cancer. We found that rationally designed synthetic CDN derivative molecules, including one with an Rp,Rp dithio diastereomer and noncanonical c[A(2',5')pA(3',5')p] phosphate bridge structure, enhanced antitumor efficacy of STINGVAX in multiple aggressive therapeutic models of established cancer in mice. Antitumor activity was STING-dependent and correlated with increased activation of dendritic cells and tumor antigen-specific CD8(+) T cells. Tumors from STINGVAX-treated mice demonstrated marked PD-L1 (programmed death ligand 1) up-regulation, which was associated with tumor-infiltrating CD8(+)IFNγ(+) T cells. When combined with PD-1 (programmed death 1) blockade, STINGVAX induced regression of palpable, poorly immunogenic tumors that did not respond to PD-1 blockade alone.
Copyright © 2015, American Association for the Advancement of Science.
Conflict of interest statement
Figures
References
-
- Morgan RA, Dudley ME, Wunderlich JR, Hughes MS, Yang JC, Sherry RM, Royal RE, Topalian SL, Kammula US, Restifo NP, Zheng Z, Nahvi A, de Vries CR, Rogers-Freezer LJ, Mavroukakis SA, Rosenberg SA. Cancer regression in patients after transfer of genetically engineered lymphocytes. Science. 2006;314:126–129. - PMC - PubMed
-
- Scholler J, Brady TL, Binder-Scholl G, Hwang WT, Plesa G, Hege KM, Vogel AN, Kalos M, Riley JL, Deeks SG, Mitsuyasu RT, Bernstein WB, Aronson NE, Levine BL, Bushman FD, June CH. Decade-long safety and function of retroviral-modified chimeric antigen receptor T cells. Sci Transl Med. 2012;4:132ra53. - PMC - PubMed
-
- Steinman RM. Decisions about dendritic cells: Past, present, and future. Annu Rev Immunol. 2012;30:1–22. - PubMed
-
- Mellman I. Dendritic cells: Master regulators of the immune response. Cancer Immunol Res. 2013;1:145–149. - PubMed
-
- Dubensky TW, Jr, Reed SG. Adjuvants for cancer vaccines. Semin Immunol. 2010;22:155–161. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
