

Origin of thoracic spinal network activity during locomotor-like activity in the neonatal rat
- PMID: 25878284
- PMCID: PMC6605171
- DOI: 10.1523/JNEUROSCI.4145-14.2015
Origin of thoracic spinal network activity during locomotor-like activity in the neonatal rat
Abstract
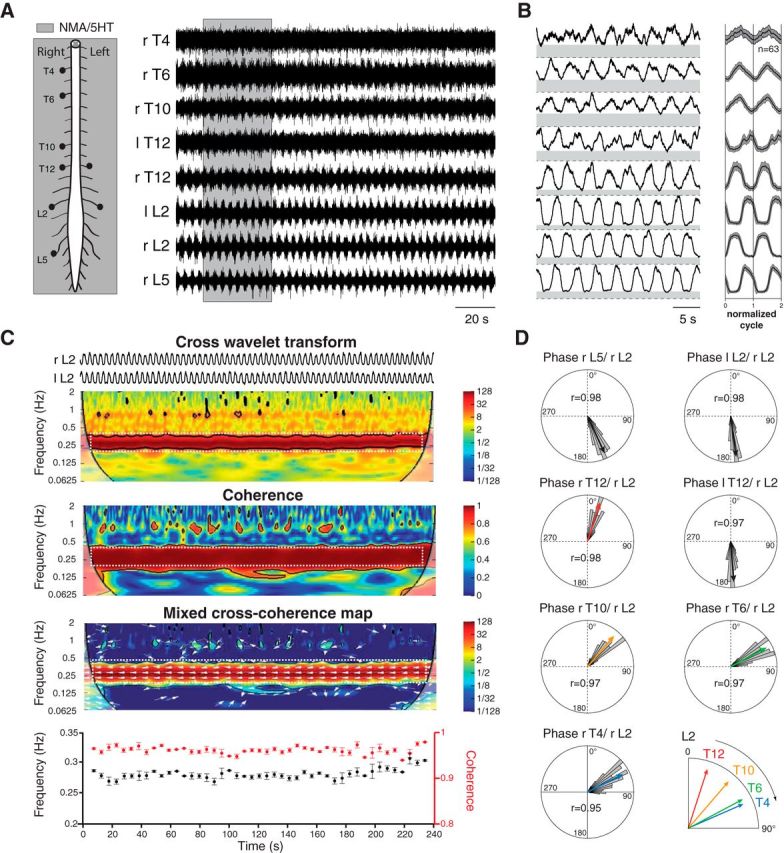
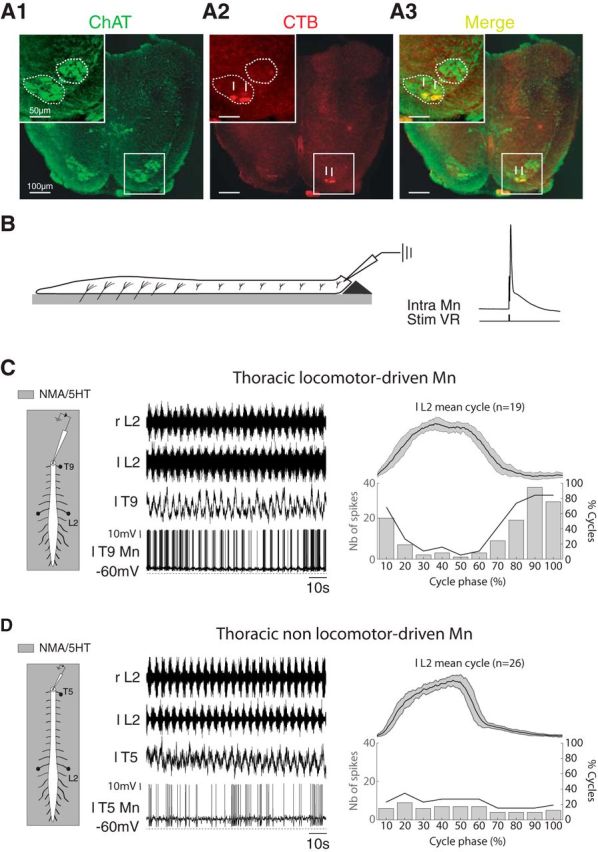
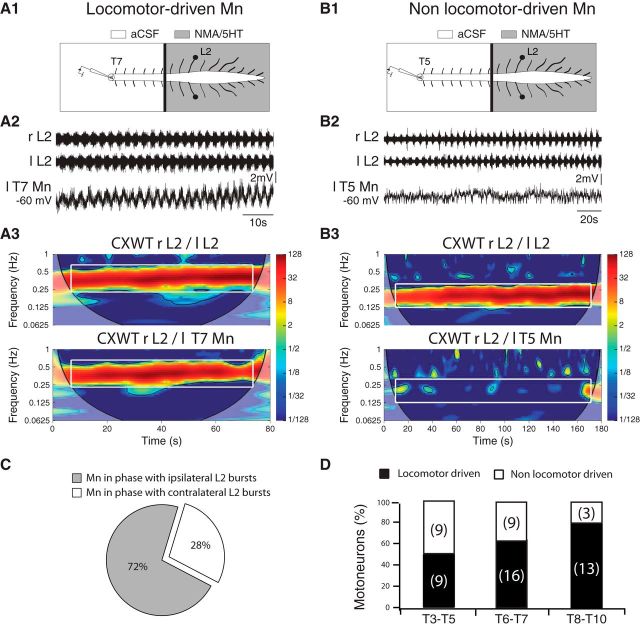
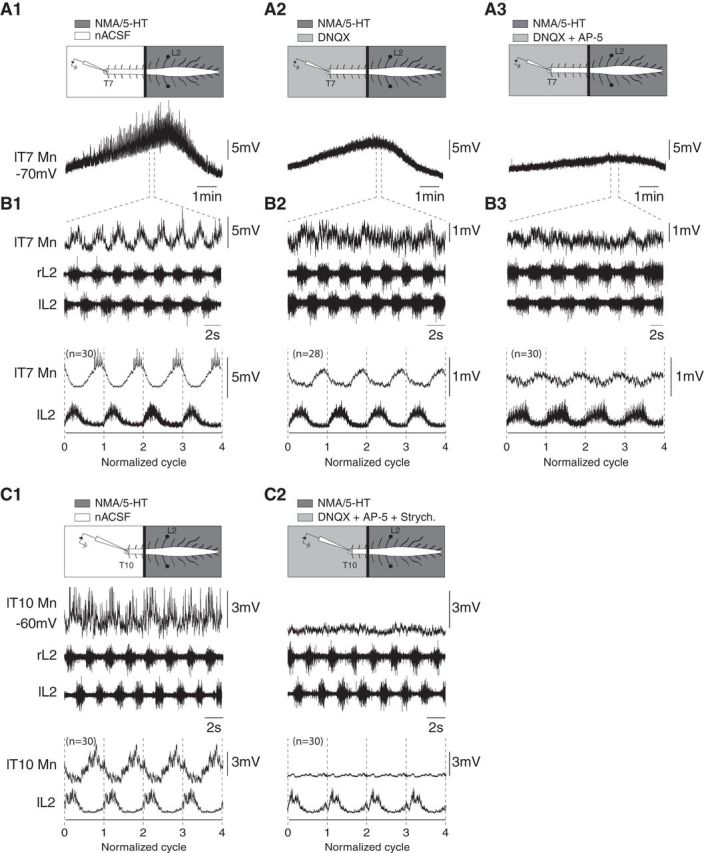
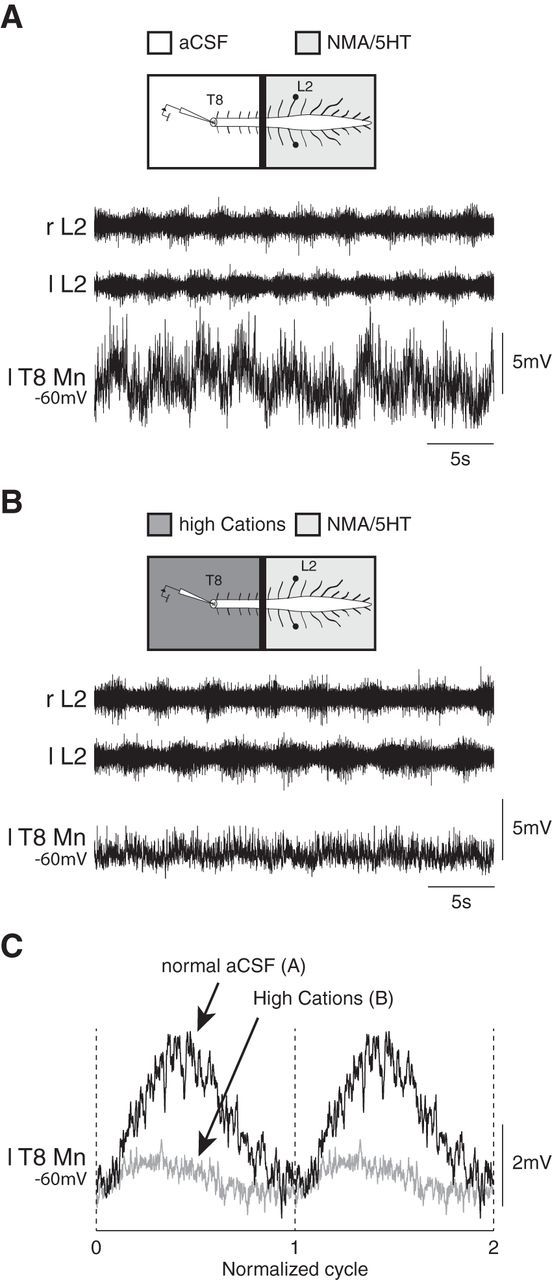
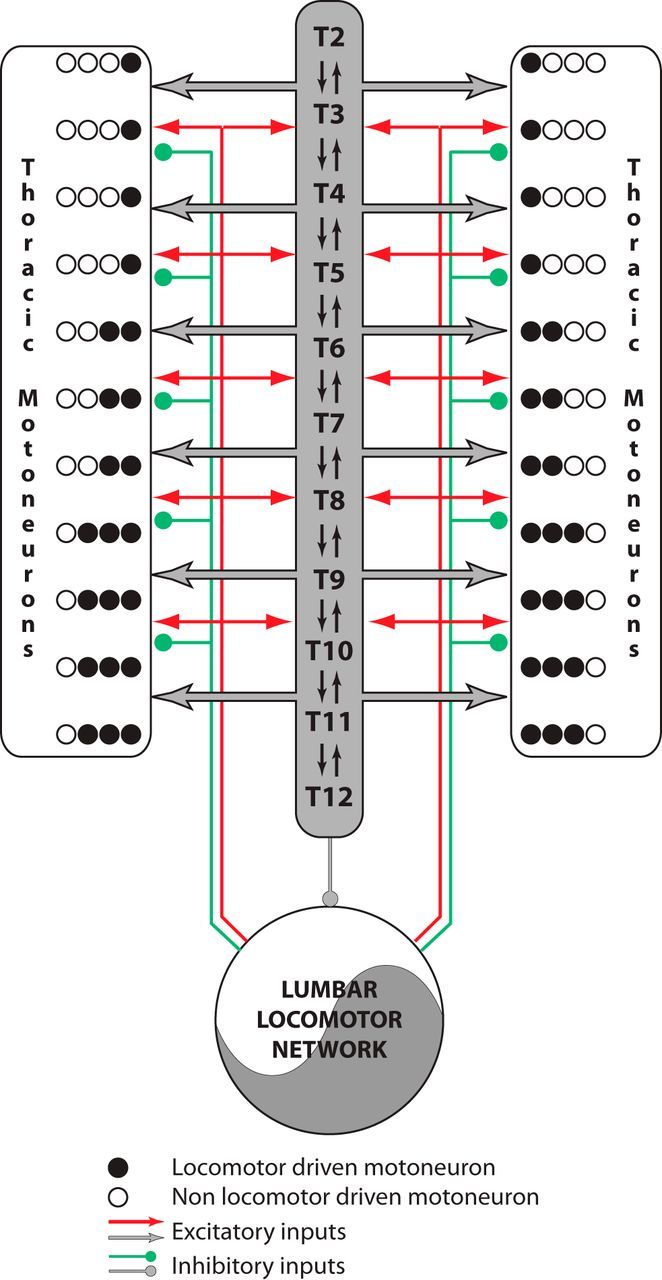
Effective quadrupedal locomotor behaviors require the coordination of many muscles in the limbs, back, neck, and tail. Because of the spinal motoneuronal somatotopic organization, motor coordination implies interactions among distant spinal networks. Here, we investigated some of the interactions between the lumbar locomotor networks that control limb movements and the thoracic networks that control the axial muscles involved in trunk movement. For this purpose, we used an in vitro isolated newborn rat spinal cord (from T2 to sacrococcygeal) preparation. Using extracellular ventral root recordings, we showed that, while the thoracic cord possesses an intrinsic rhythmogenic capacity, the lumbar circuits, if they are rhythmically active, will entrain the rhythmicity of the thoracic circuitry. However, if the lumbar circuits are rhythmically active, these latter circuits will entrain the rhythmicity of the thoracic circuitry. Blocking the synaptic transmission in some thoracic areas revealed that the lumbar locomotor network could trigger locomotor bursting in distant thoracic segments through short and long propriospinal pathways. Patch-clamp recordings revealed that 72% of the thoracic motoneurons (locomotor-driven motoneurons) expressed membrane potential oscillations and spiking activity coordinated with the locomotor activity expressed by the lumbar cord. A biphasic excitatory (glutamatergic)/inhibitory (glycinergic) synaptic drive was recorded in thoracic locomotor-driven motoneurons. Finally, we found evidence that part of this locomotor drive involved a monosynaptic component coming directly from the lumbar locomotor network. We conclude that the lumbar locomotor network plays a central role in the generation of locomotor outputs in the thoracic cord by acting at both the premotoneuronal and motoneuronal levels.
Keywords: axial; coordination; in vitro; locomotion; posture; spinal cord.
Copyright © 2015 the authors 0270-6474/15/356117-14$15.00/0.
Figures
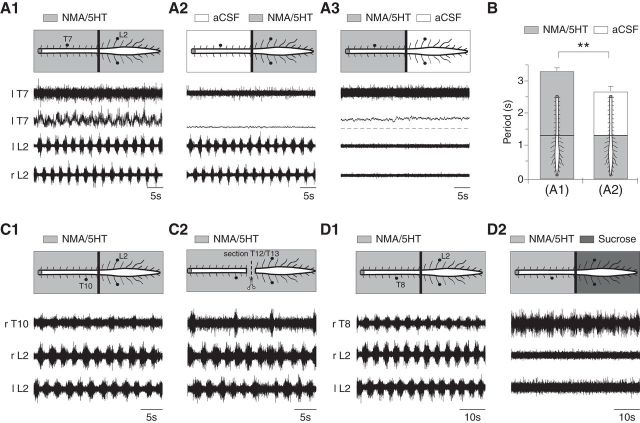
 ) extracellular activities were recorded in whole spinal cords (A1) or lumbar (A2) or thoracic (A3) segments treated with NMA (15 μ
) extracellular activities were recorded in whole spinal cords (A1) or lumbar (A2) or thoracic (A3) segments treated with NMA (15 μ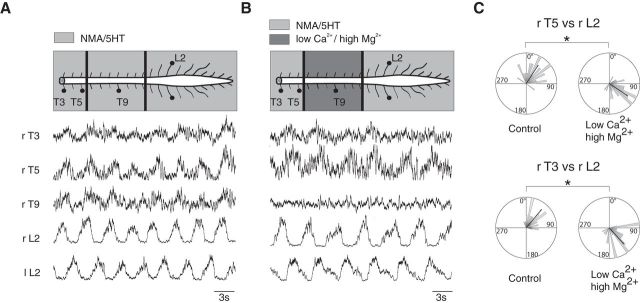
References
-
- Berens P. CircStat: a Matlab toolbox for circular statistics. J Statistical Software. 2009;31:1–21.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources