

Target of Rapamycin Complex 2 Regulates Actin Polarization and Endocytosis via Multiple Pathways
- PMID: 25882841
- PMCID: PMC4463442
- DOI: 10.1074/jbc.M114.627794
Target of Rapamycin Complex 2 Regulates Actin Polarization and Endocytosis via Multiple Pathways
Abstract
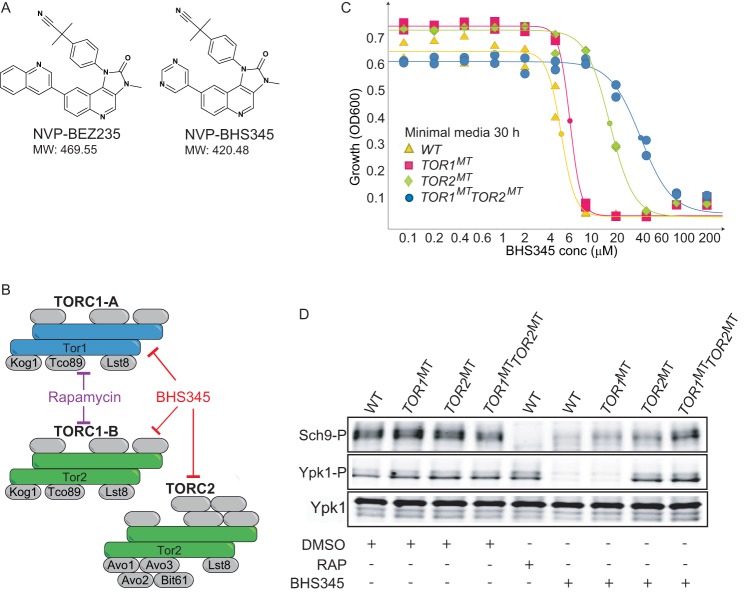
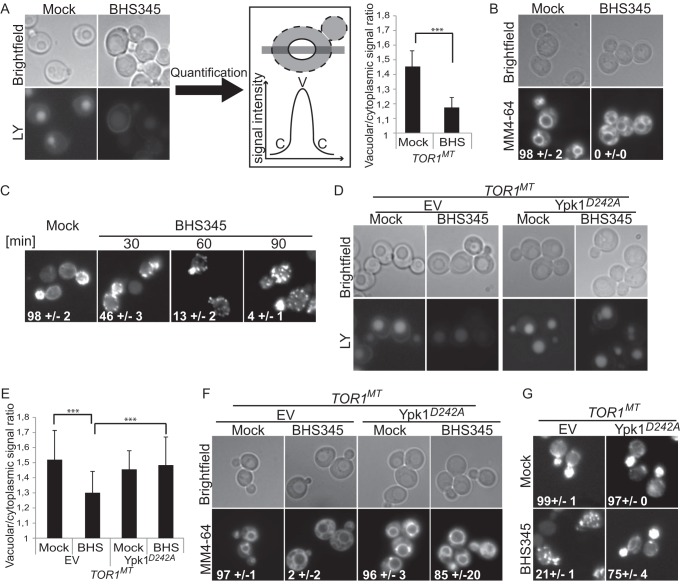
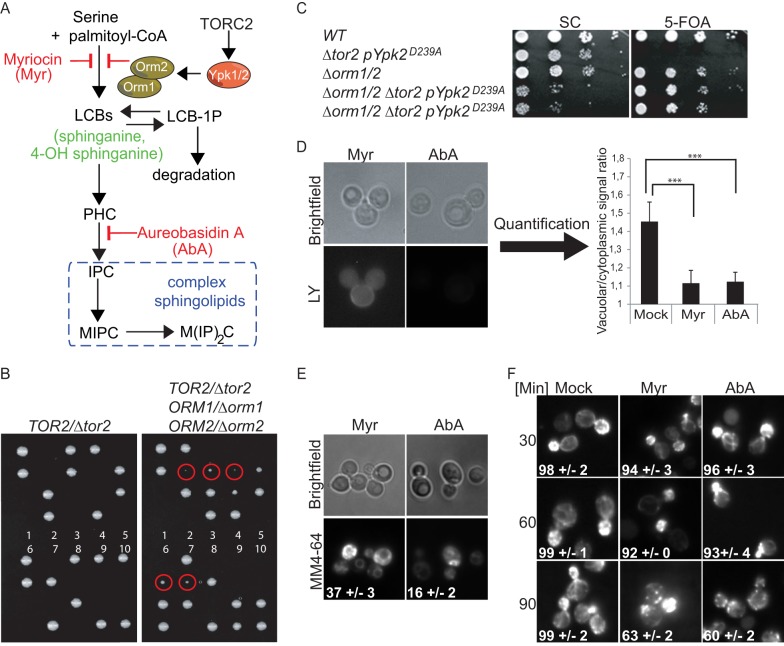
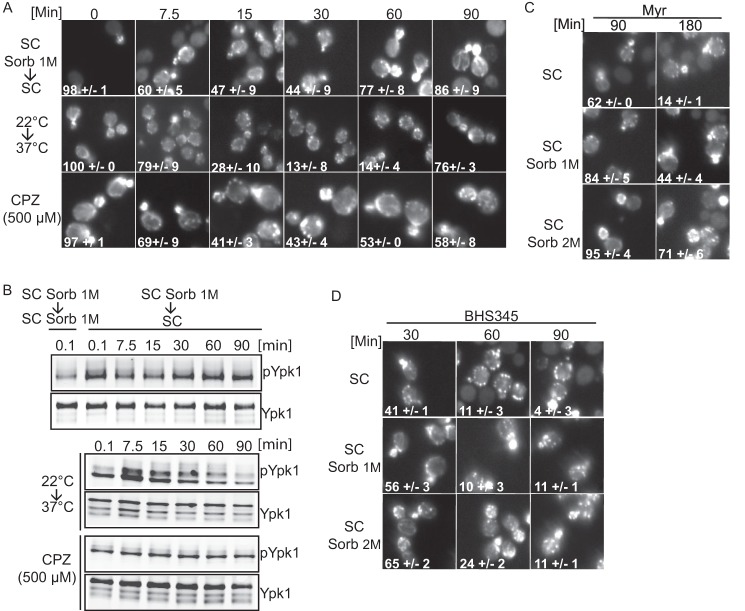
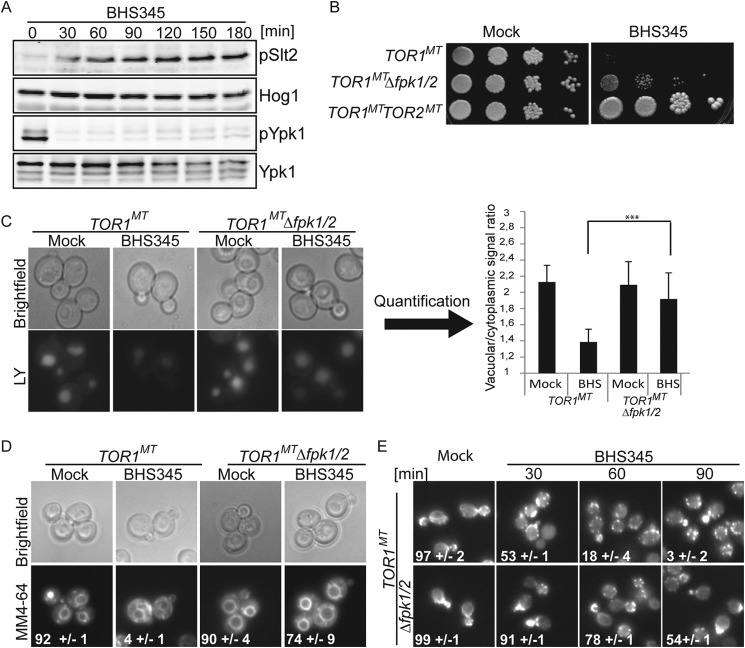
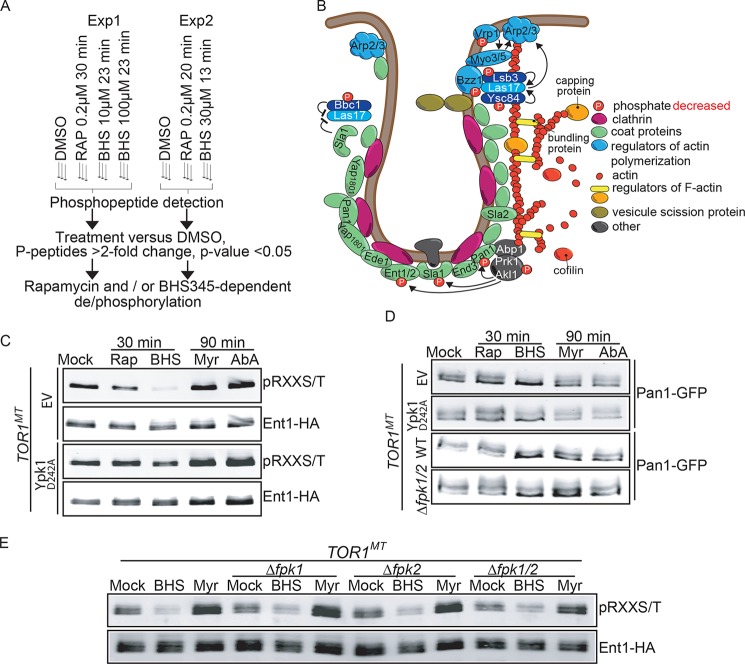
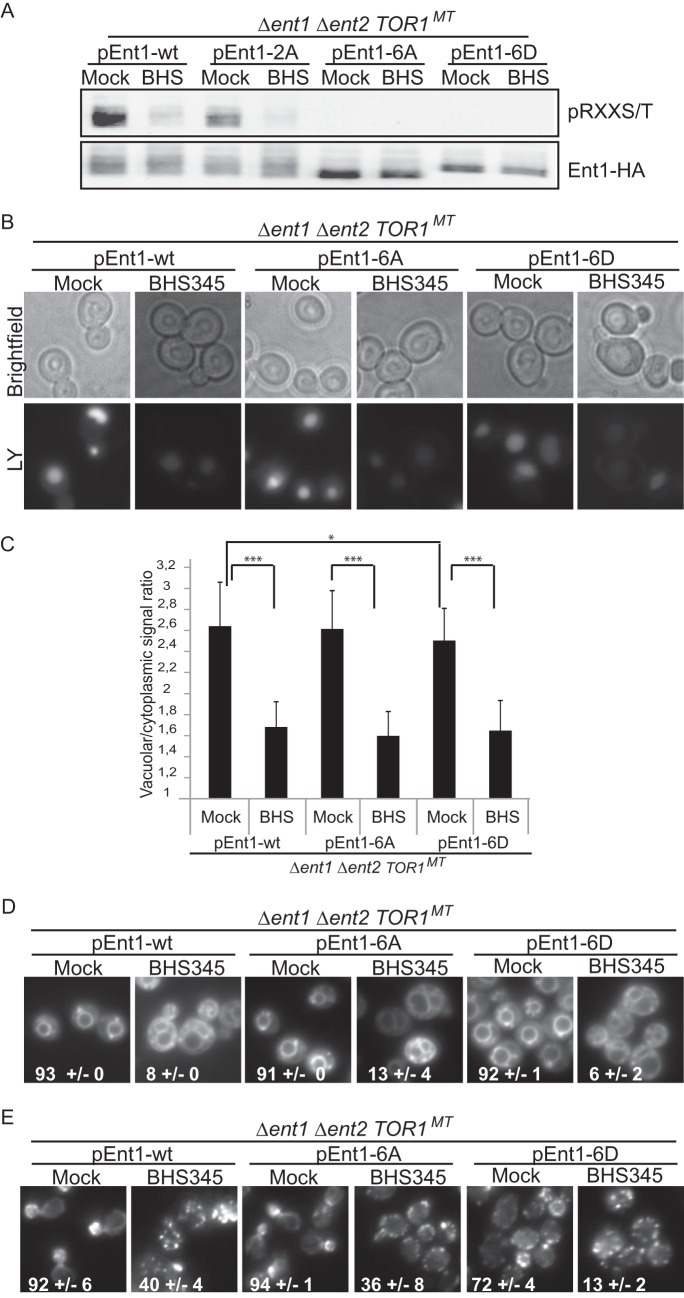
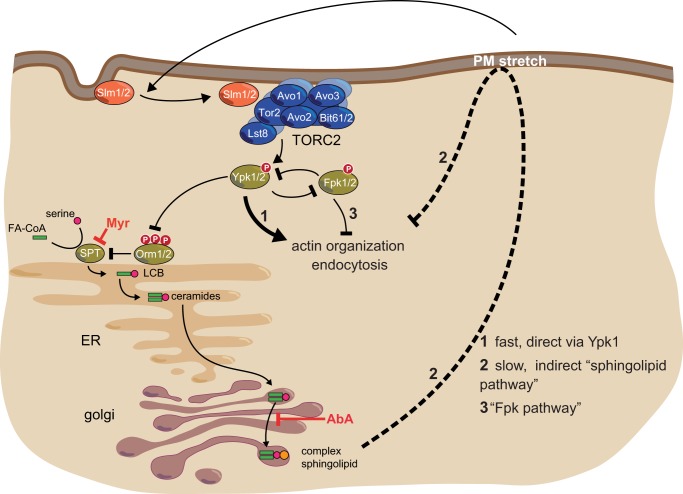
Target of rapamycin is a Ser/Thr kinase that operates in two conserved multiprotein complexes, TORC1 and TORC2. Unlike TORC1, TORC2 is insensitive to rapamycin, and its functional characterization is less advanced. Previous genetic studies demonstrated that TORC2 depletion leads to loss of actin polarization and loss of endocytosis. To determine how TORC2 regulates these readouts, we engineered a yeast strain in which TORC2 can be specifically and acutely inhibited by the imidazoquinoline NVP-BHS345. Kinetic analyses following inhibition of TORC2, supported with quantitative phosphoproteomics, revealed that TORC2 regulates these readouts via distinct pathways as follows: rapidly through direct protein phosphorylation cascades and slowly through indirect changes in the tensile properties of the plasma membrane. The rapid signaling events are mediated in large part through the phospholipid flippase kinases Fpk1 and Fpk2, whereas the slow signaling pathway involves increased plasma membrane tension resulting from a gradual depletion of sphingolipids. Additional hits in our phosphoproteomic screens highlight the intricate control TORC2 exerts over diverse aspects of eukaryote cell physiology.
Keywords: cell signaling; lipid signaling; phosphoproteome; plasma membrane; protein kinase; target of rapamycin (TOR); target of rapamycin complex 2, Ypk1, sphingolipids, actin polarization, endocytosis, flippase kinase.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Cafferkey R., Young P. R., McLaughlin M. M., Bergsma D. J., Koltin Y., Sathe G. M., Faucette L., Eng W. K., Johnson R. K., Livi G. P. (1993) Dominant missense mutations in a novel yeast protein related to mammalian phosphatidylinositol 3-kinase and VPS34 abrogate rapamycin cytotoxicity. Mol. Cell. Biol. 13, 6012–6023 - PMC - PubMed
-
- Heitman J., Movva N. R., Hall M. N. (1991) Targets for cell cycle arrest by the immunosuppressant rapamycin in yeast. Science 253, 905–909 - PubMed
-
- Kunz J., Henriquez R., Schneider U., Deuter-Reinhard M., Movva N. R., Hall M. N. (1993) Target of rapamycin in yeast, TOR2, is an essential phosphatidylinositol kinase homolog required for G1 progression. Cell 73, 585–596 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
