

CRL3IBTK Regulates the Tumor Suppressor Pdcd4 through Ubiquitylation Coupled to Proteasomal Degradation
- PMID: 25882842
- PMCID: PMC4447969
- DOI: 10.1074/jbc.M114.634535
CRL3IBTK Regulates the Tumor Suppressor Pdcd4 through Ubiquitylation Coupled to Proteasomal Degradation
Abstract
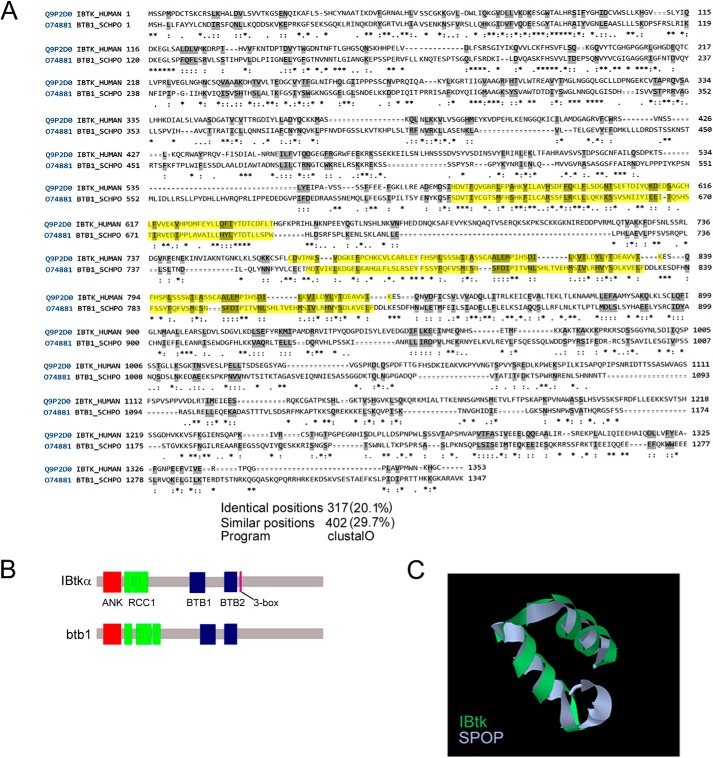
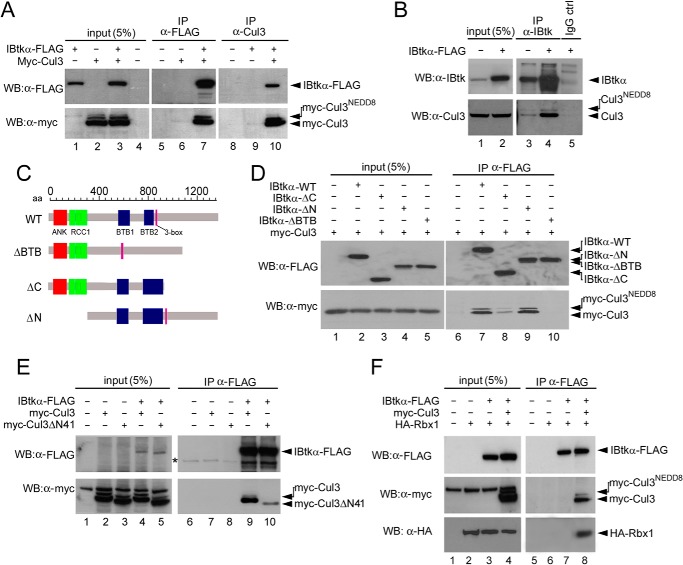
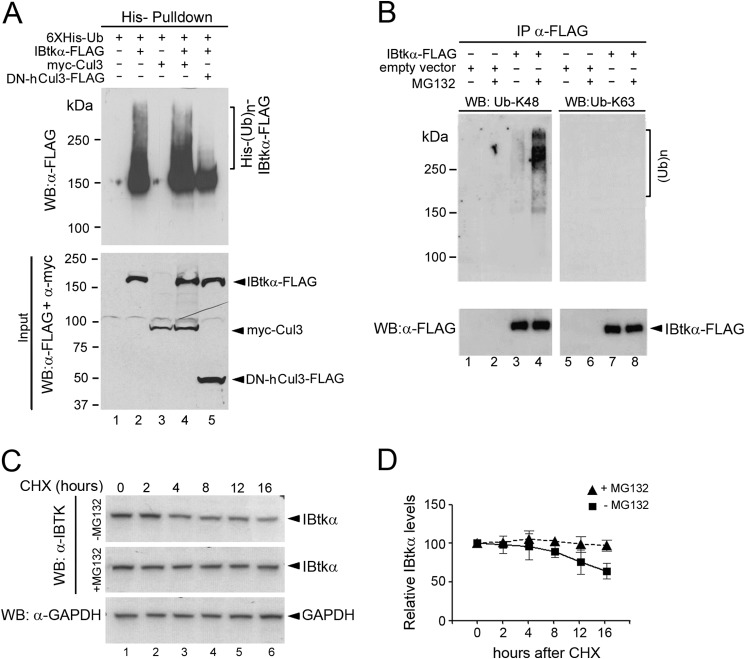
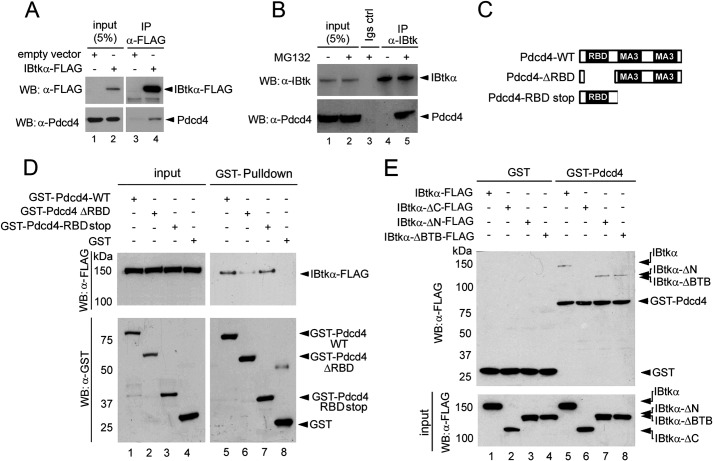
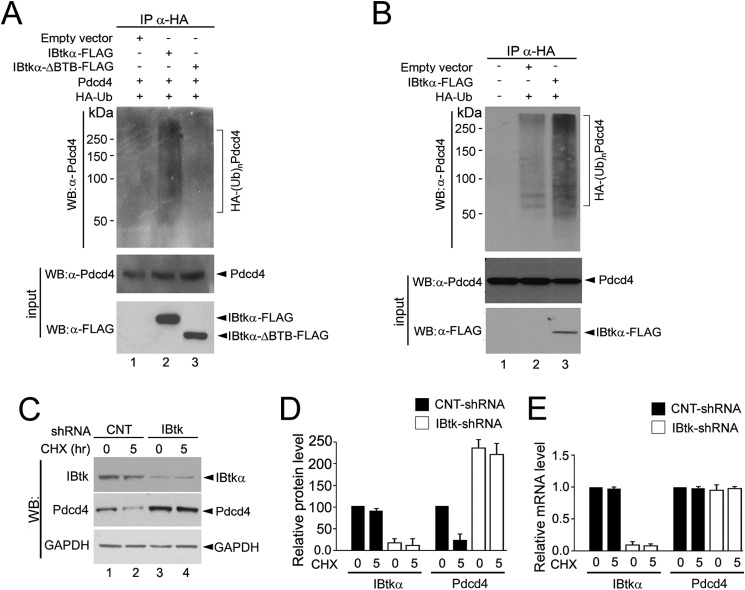
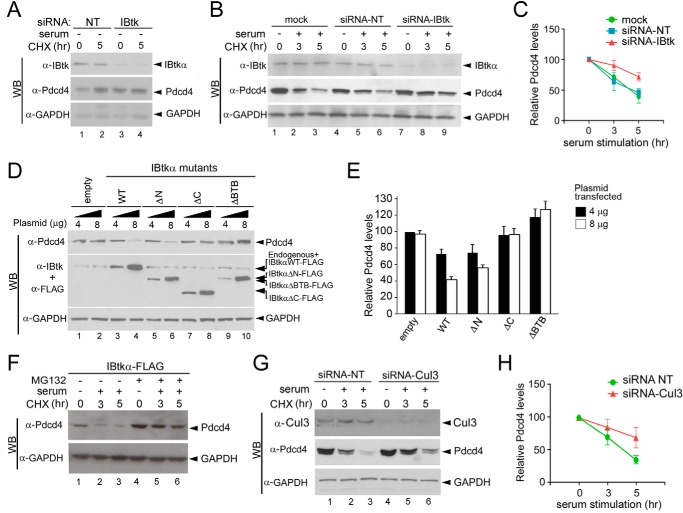
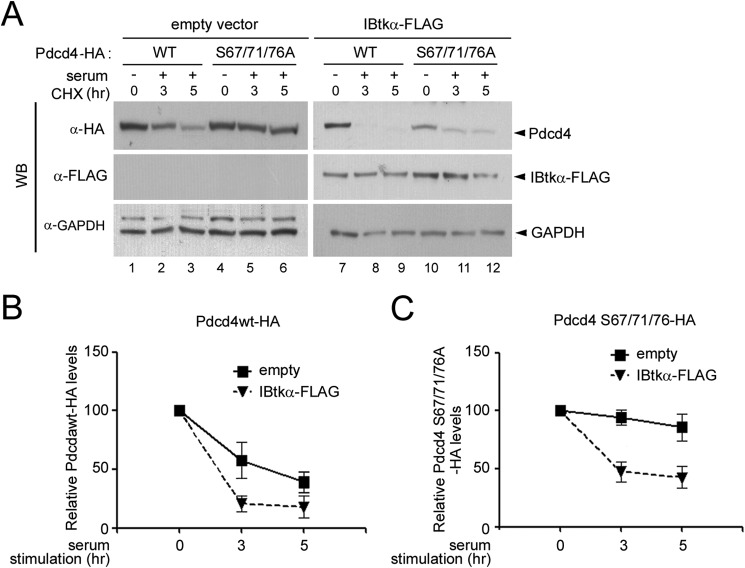
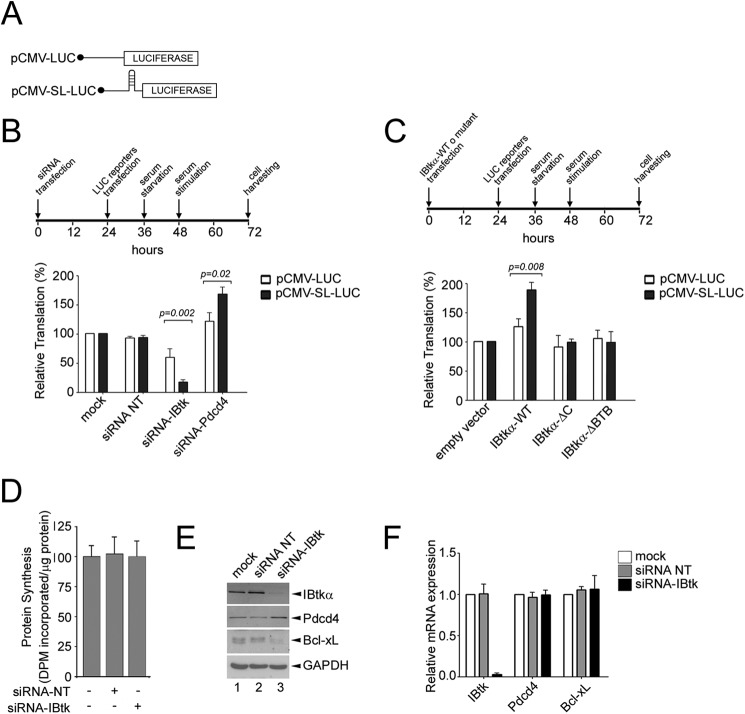
The human inhibitor of Bruton's tyrosine kinase isoform α (IBtkα) is a BTB protein encoded by the IBTK gene, which maps to chromosomal locus 6q14.1, a mutational hot spot in lymphoproliferative disorders. Here, we demonstrate that IBtkα forms a CRL3(IBTK) complex promoting its self-ubiquitylation. We identified the tumor suppressor Pdcd4 as IBtkα interactor and ubiquitylation substrate of CRL3(IBTK) for proteasomal degradation. Serum-induced degradation of Pdcd4 required both IBtkα and Cul3, indicating that CRL3(IBTK) regulated the Pdcd4 stability in serum signaling. By promoting Pdcd4 degradation, IBtkα counteracted the suppressive effect of Pdcd4 on translation of reporter luciferase mRNAs with stem-loop structured or unstructured 5'-UTR. IBtkα depletion by RNAi caused Pdcd4 accumulation and decreased the translation of Bcl-xL mRNA, a well known target of Pdcd4 repression. By characterizing CRL3(IBTK) as a novel ubiquitin ligase, this study provides new insights into regulatory mechanisms of cellular pathways, such as the Pdcd4-dependent translation of mRNAs.
Keywords: E3 ubiquitin ligase; IBtk; Pdcd4; serum; translation regulation; tumor suppressor gene; ubiquitylation (ubiquitination).
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Hershko A., Ciechanover A. (1998) The ubiquitin system. Annu. Rev. Biochem. 67, 425–479 - PubMed
-
- Pickart C. M. (2004) Back to the future with ubiquitin. Cell 116, 181–190 - PubMed
-
- Petroski M. D., Deshaies R. J. (2005) Function and regulation of cullin-RING ubiquitin ligases. Nat. Rev. Mol. Cell Biol. 6, 9–20 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
