

Sequence analysis reveals a conserved extension in the capping enzyme of the alphavirus supergroup, and a homologous domain in nodaviruses
- PMID: 25886938
- PMCID: PMC4392871
- DOI: 10.1186/s13062-015-0050-0
Sequence analysis reveals a conserved extension in the capping enzyme of the alphavirus supergroup, and a homologous domain in nodaviruses
Abstract
Background: Members of the alphavirus supergroup include human pathogens such as chikungunya virus, hepatitis E virus and rubella virus. They encode a capping enzyme with methyltransferase-guanylyltransferase (MTase-GTase) activity, which is an attractive drug target owing to its unique mechanism. However, its experimental study has proven very difficult.
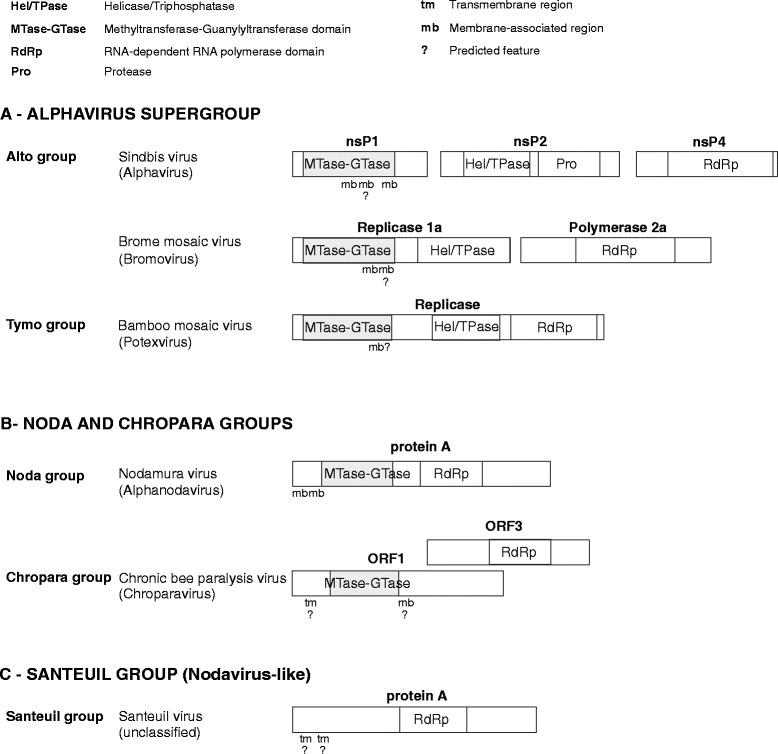
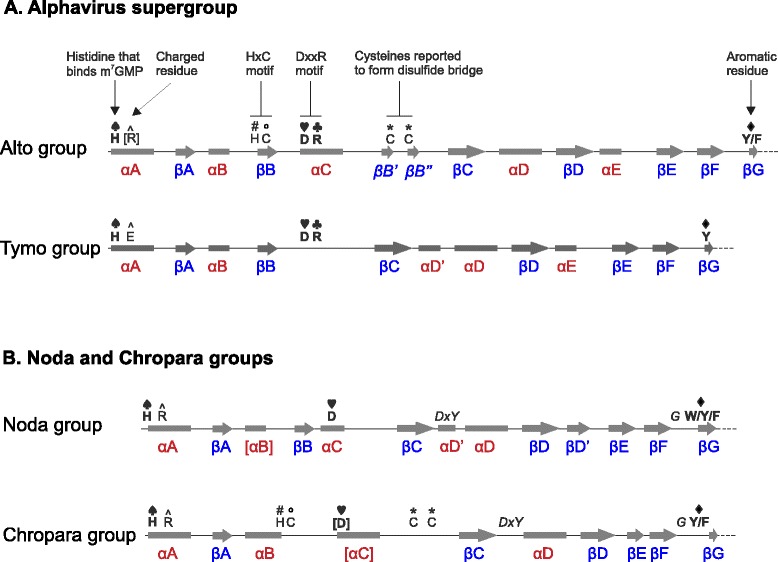
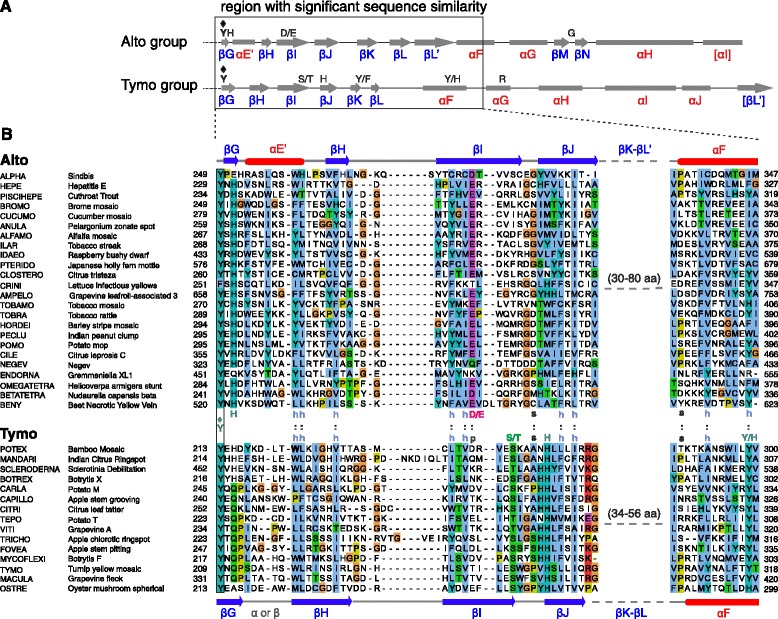
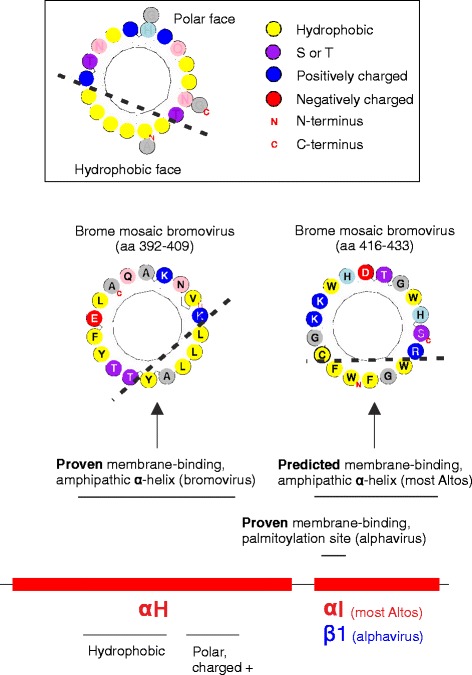
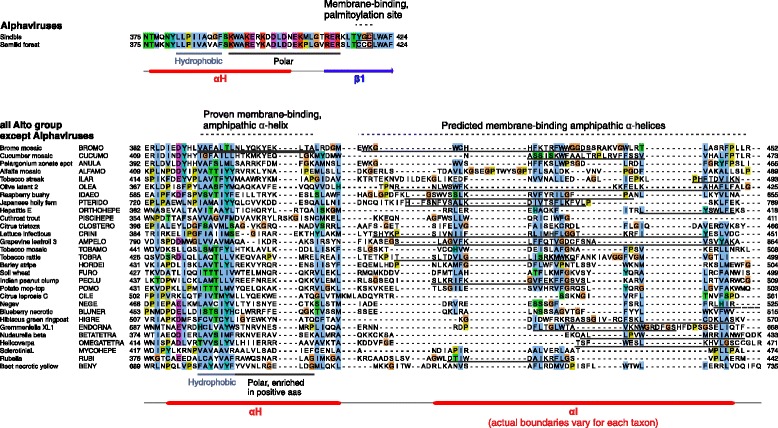
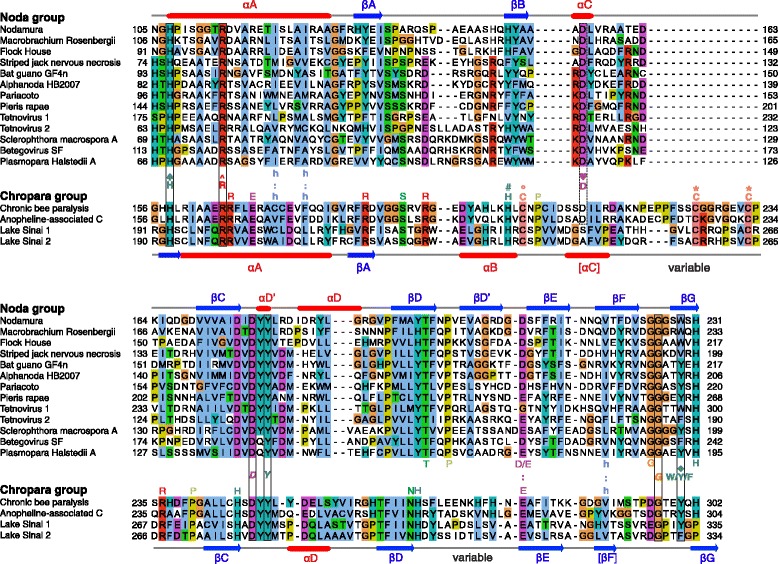
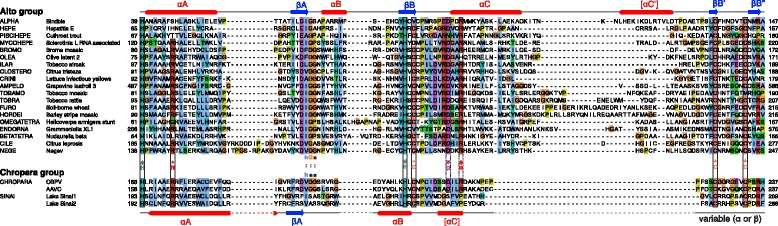
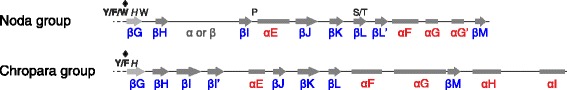
Results: We examined over 50 genera of viruses by sequence analyses. Earlier studies showed that the MTase-GTase contains a "Core" region conserved in sequence. We show that it is followed by a long extension, which we termed "Iceberg" region, whose secondary structure, but not sequence, is strikingly conserved throughout the alphavirus supergroup. Sequence analyses strongly suggest that the minimal capping domain corresponds to the Core and Iceberg regions combined, which is supported by earlier experimental data. The Iceberg region contains all known membrane association sites that contribute to the assembly of viral replication factories. We predict that it may also contain an overlooked, widely conserved membrane-binding amphipathic helix. Unexpectedly, we detected a sequence homolog of the alphavirus MTase-GTase in taxa related to nodaviruses and to chronic bee paralysis virus. The presence of a capping enzyme in nodaviruses is biologically consistent, since they have capped genomes but replicate in the cytoplasm, where no cellular capping enzyme is present. The putative MTase-GTase domain of nodaviruses also contains membrane-binding sites that may drive the assembly of viral replication factories, revealing an unsuspected parallel with the alphavirus supergroup.
Conclusions: Our work will guide the functional analysis of the alphaviral MTase-GTase and the production of domains for structure determination. The identification of a homologous domain in a simple model system, nodaviruses, which replicate in numerous eukaryotic cell systems (yeast, flies, worms, mammals, and plants), can further help crack the function and structure of the enzyme.
Figures
References
-
- Goldbach R. Genome similarities between plant and animal RNA viruses. Microbiol Sci. 1987;4(7):197–202. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
