

Tau Trimers Are the Minimal Propagation Unit Spontaneously Internalized to Seed Intracellular Aggregation
- PMID: 25887395
- PMCID: PMC4463437
- DOI: 10.1074/jbc.M115.652693
Tau Trimers Are the Minimal Propagation Unit Spontaneously Internalized to Seed Intracellular Aggregation
Abstract
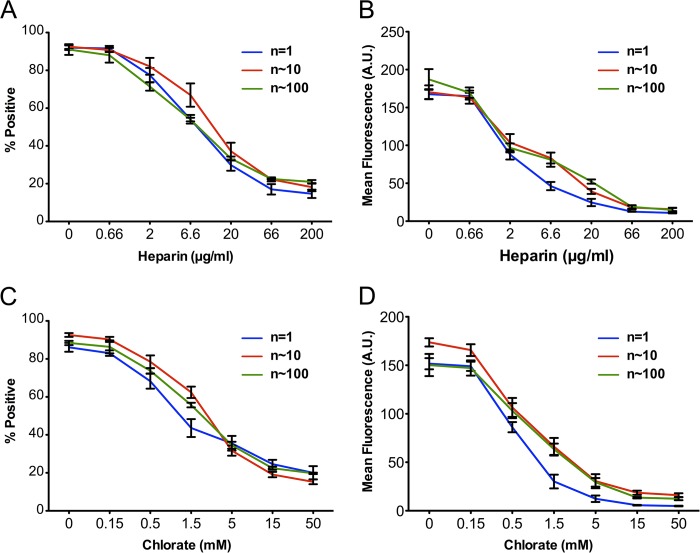
Tau amyloid assemblies propagate aggregation from the outside to the inside of a cell, which may mediate progression of the tauopathies. The critical size of Tau assemblies, or "seeds," responsible for this activity is currently unknown, but this could be important for the design of effective therapies. We studied recombinant Tau repeat domain (RD) and Tau assemblies purified from Alzheimer disease (AD) brain composed largely of full-length Tau. Large RD fibrils were first sonicated to create a range of assembly sizes. We confirmed our ability to resolve stable assemblies ranging from n = 1 to >100 units of Tau using size exclusion chromatography, fluorescence correlation spectroscopy, cross-linking followed by Western blot, and mass spectrometry. All recombinant Tau assemblies bound heparan sulfate proteoglycans on the cell surface, which are required for Tau uptake and seeding, because they were equivalently sensitive to inhibition by heparin and chlorate. However, cells only internalized RD assemblies of n ≥ 3 units. We next analyzed Tau assemblies from AD or control brains. AD brains contained aggregated species, whereas normal brains had predominantly monomer, and no evidence of large assemblies. HEK293 cells and primary neurons spontaneously internalized Tau of n ≥ 3 units from AD brain in a heparin- and chlorate-sensitive manner. Only n ≥ 3-unit assemblies from AD brain spontaneously seeded intracellular Tau aggregation in HEK293 cells. These results indicate that a clear minimum size (n = 3) of Tau seed exists for spontaneous propagation of Tau aggregation from the outside to the inside of a cell, whereas many larger sizes of soluble aggregates trigger uptake and seeding.
Keywords: Tau protein; amyloid; endocytosis; neurodegenerative disease; prion; structure.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures







References
-
- Lee V. M.-Y., Goedert M., Trojanowski J. Q. (2001) Neurodegenerative tauopathies. Annu. Rev. Neurosci. 24, 1121–1159 - PubMed
-
- de Calignon A. A., Polydoro M. M., Suárez-Calvet M., William C., Adamowicz D. H. D., Kopeikina K. J. K., Pitstick R., Sahara N., Ashe K. H. K., Carlson G. A., Spires-Jones T. L. T., Hyman B. T. (2012) Propagation of tau pathology in a model of early Alzheimer's disease. Neuron 73, 685–697 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
