

Pioneer transcription factors target partial DNA motifs on nucleosomes to initiate reprogramming
- PMID: 25892221
- PMCID: PMC4409934
- DOI: 10.1016/j.cell.2015.03.017
Pioneer transcription factors target partial DNA motifs on nucleosomes to initiate reprogramming
Abstract
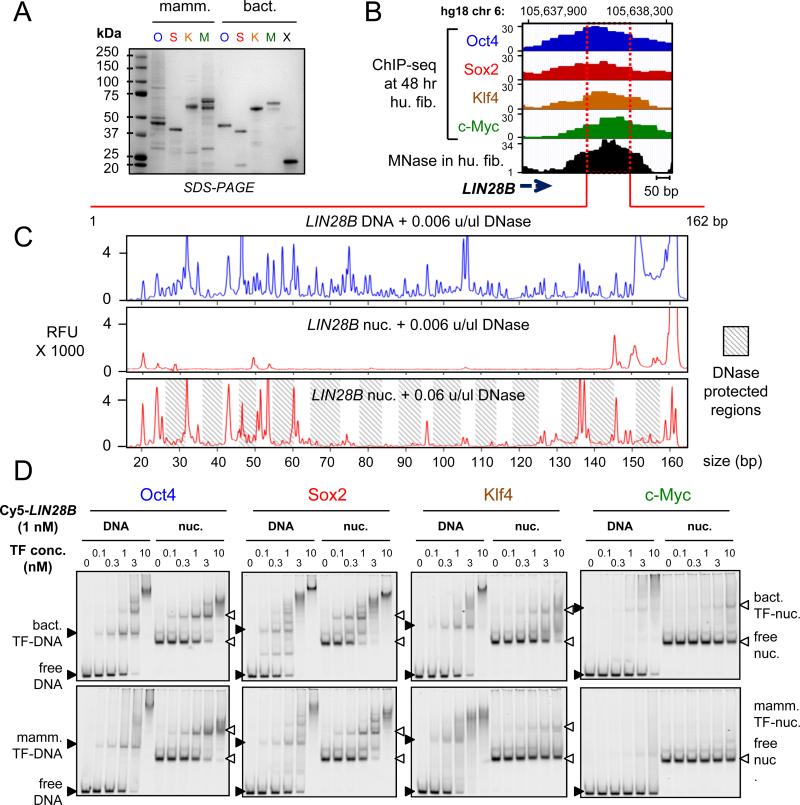
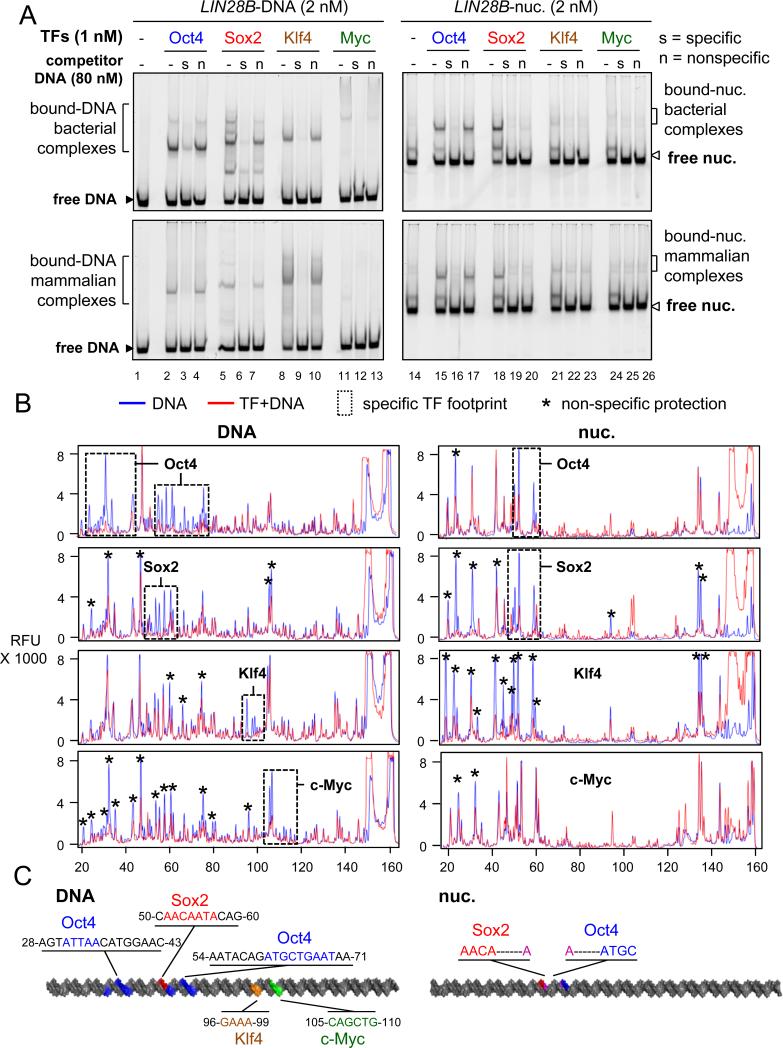
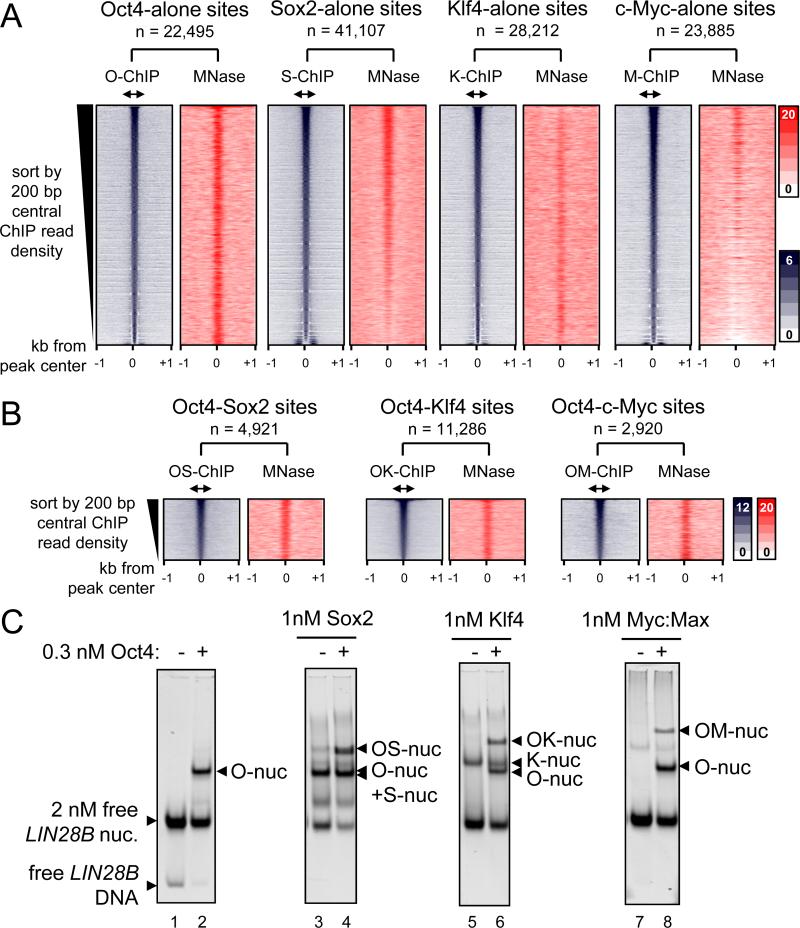
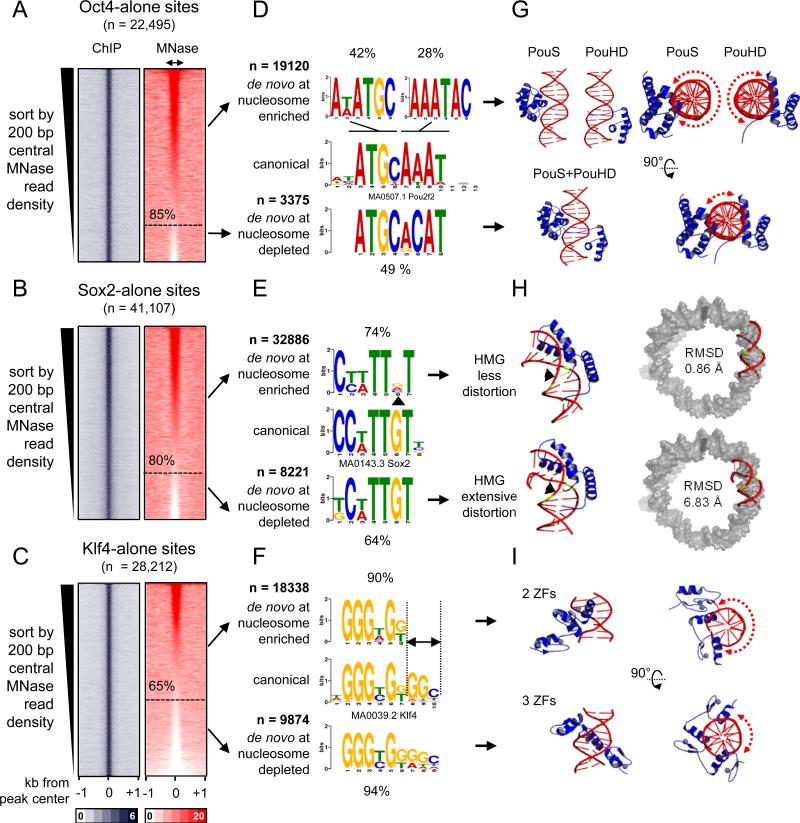
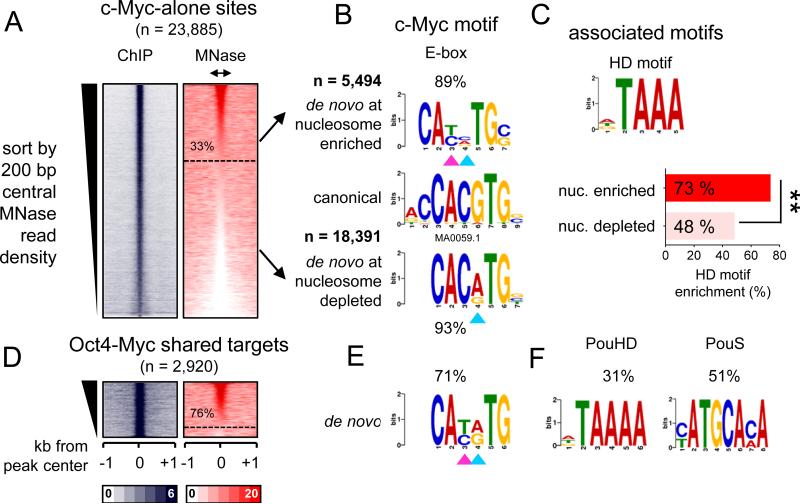
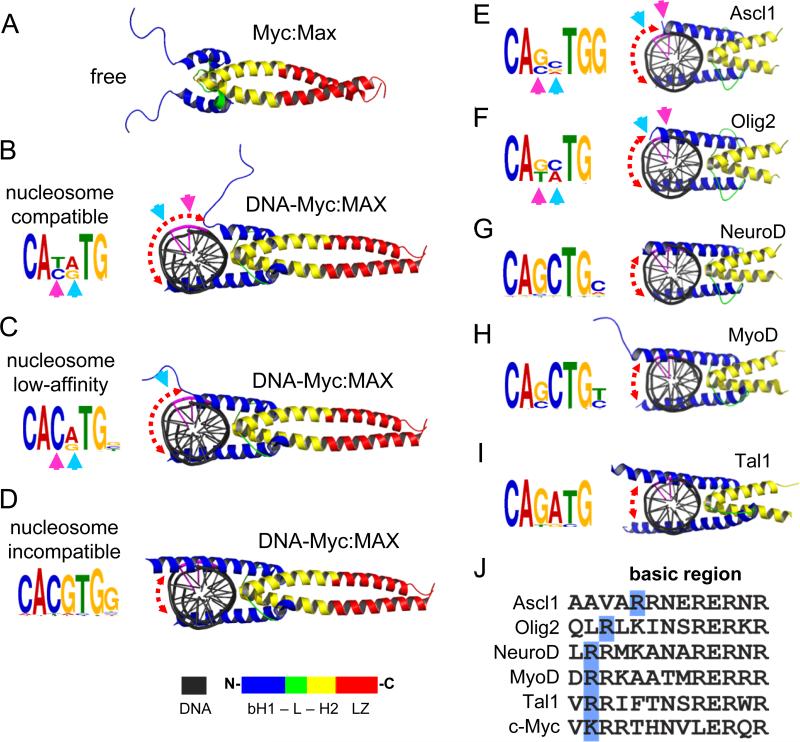
Pioneer transcription factors (TFs) access silent chromatin and initiate cell-fate changes, using diverse types of DNA binding domains (DBDs). FoxA, the paradigm pioneer TF, has a winged helix DBD that resembles linker histone and thereby binds its target sites on nucleosomes and in compacted chromatin. Herein, we compare the nucleosome and chromatin targeting activities of Oct4 (POU DBD), Sox2 (HMG box DBD), Klf4 (zinc finger DBD), and c-Myc (bHLH DBD), which together reprogram somatic cells to pluripotency. Purified Oct4, Sox2, and Klf4 proteins can bind nucleosomes in vitro, and in vivo they preferentially target silent sites enriched for nucleosomes. Pioneer activity relates simply to the ability of a given DBD to target partial motifs displayed on the nucleosome surface. Such partial motif recognition can occur by coordinate binding between factors. Our findings provide insight into how pioneer factors can target naive chromatin sites.
Copyright © 2015 Elsevier Inc. All rights reserved.
Figures
Comment in
-
How to Be a Pioneer: A One-Sided View.Trends Biochem Sci. 2015 Oct;40(10):547-548. doi: 10.1016/j.tibs.2015.08.011. Epub 2015 Sep 8. Trends Biochem Sci. 2015. PMID: 26358228 No abstract available.
References
-
- Biggin MD. Animal transcription networks as highly connected, quantitative continua. Dev. Cell. 2011;21:611–626. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
