

Mechanism and regulation of rapid telomere prophase movements in mouse meiotic chromosomes
- PMID: 25892231
- PMCID: PMC4417006
- DOI: 10.1016/j.celrep.2015.03.045
Mechanism and regulation of rapid telomere prophase movements in mouse meiotic chromosomes
Abstract
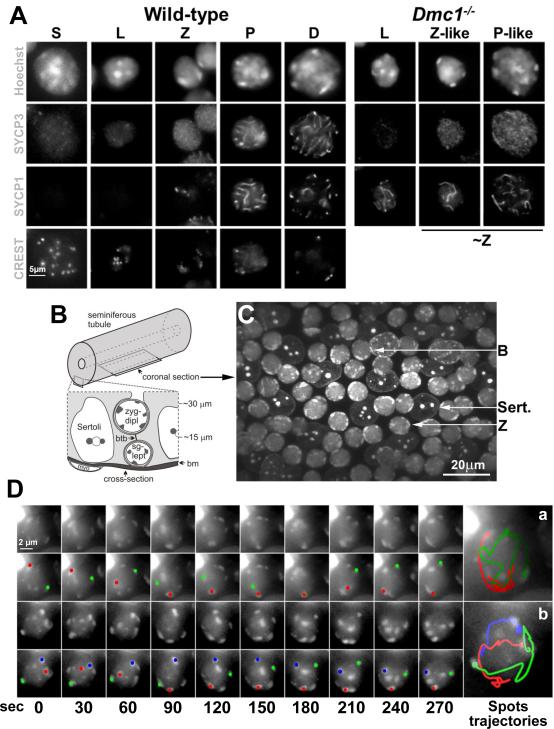
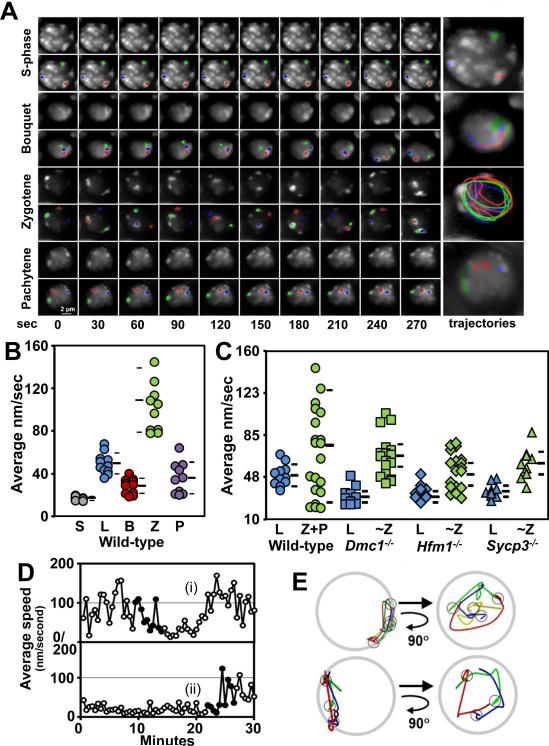
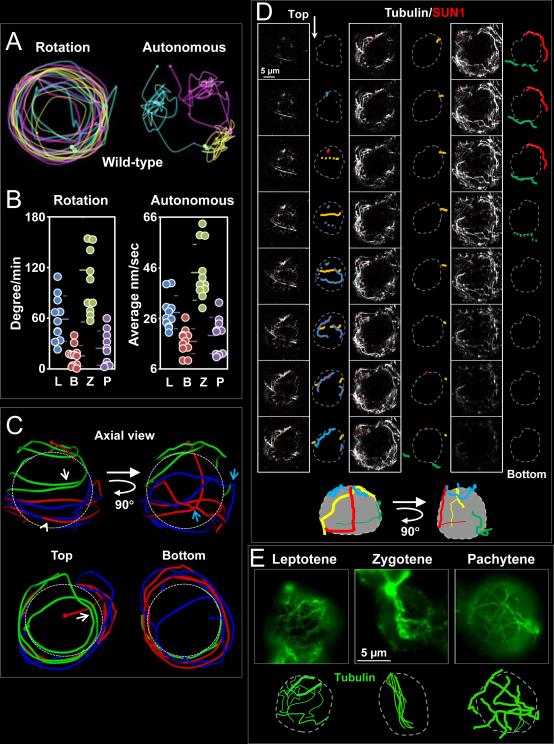
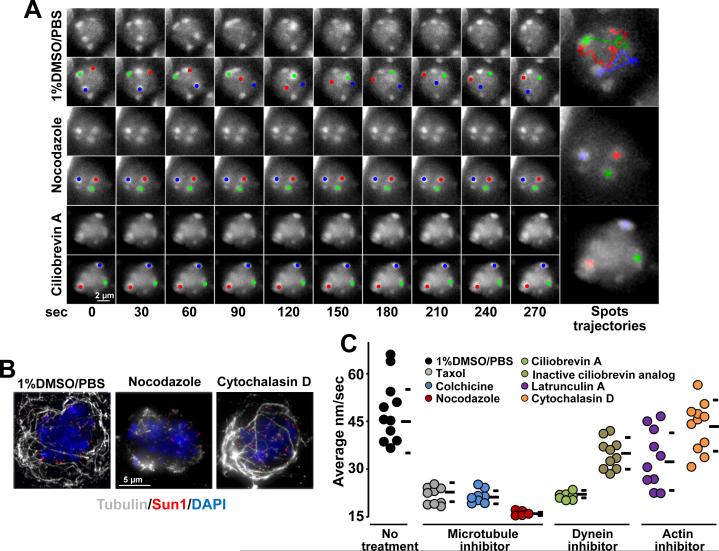
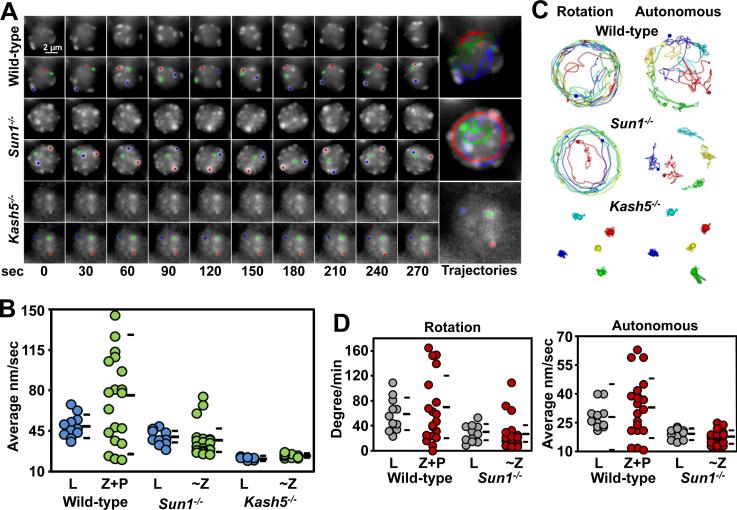
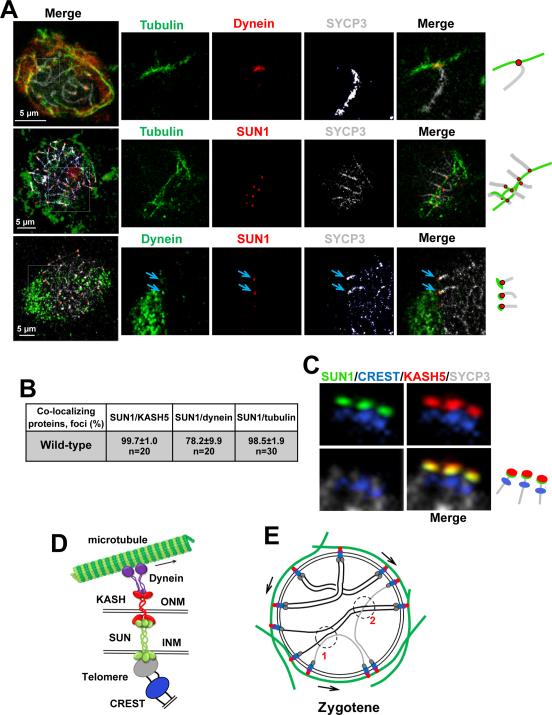
Telomere-led rapid prophase movements (RPMs) in meiotic prophase have been observed in diverse eukaryote species. A shared feature of RPMs is that the force that drives the chromosomal movements is transmitted from the cytoskeleton, through the nuclear envelope, to the telomeres. Studies in mice suggested that dynein movement along microtubules is transmitted to telomeres through SUN1/KASH5 nuclear envelope bridges to generate RPMs. We monitored RPMs in mouse seminiferous tubules using 4D fluorescence imaging and quantitative motion analysis to characterize patterns of movement in the RPM process. We find that RPMs reflect a combination of nuclear rotation and individual chromosome movements. The telomeres move along microtubule tracks that are apparently continuous with the cytoskeletal network and exhibit characteristic arrangements at different stages of prophase. Quantitative measurements confirmed that SUN1/KASH5, microtubules, and dynein, but not actin, were necessary for RPMs and that defects in meiotic recombination and synapsis resulted in altered RPMs.
Copyright © 2015 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
References
-
- Chikashige Y, Ding DQ, Funabiki H, Haraguchi T, Mashiko S, Yanagida M, Hiraoka Y. Telomere-led premeiotic chromosome movement in fission yeast. Science. 1994;264:270–273. - PubMed
-
- Chikashige Y, Haraguchi T, Hiraoka Y. Another way to move chromosomes. Chromosoma. 2007;116:497–505. - PubMed
-
- Conrad MN, Lee CY, Chao G, Shinohara M, Kosaka H, Shinohara A, Conchello JA, Dresser ME. Rapid telomere movement in meiotic prophase is promoted by NDJ1, MPS3, and CSM4 and is modulated by recombination. Cell. 2008;133:1175–1187. - PubMed
-
- Ding DQ, Chikashige Y, Haraguchi T, Hiraoka Y. Oscillatory nuclear movement in fission yeast meiotic prophase is driven by astral microtubules, as revealed by continuous observation of chromosomes and microtubules in living cells. J Cell Sci. 1998;111(Pt 6):701–712. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
