

Anisotropic stress orients remodelling of mammalian limb bud ectoderm
- PMID: 25893915
- PMCID: PMC4955842
- DOI: 10.1038/ncb3156
Anisotropic stress orients remodelling of mammalian limb bud ectoderm
Abstract
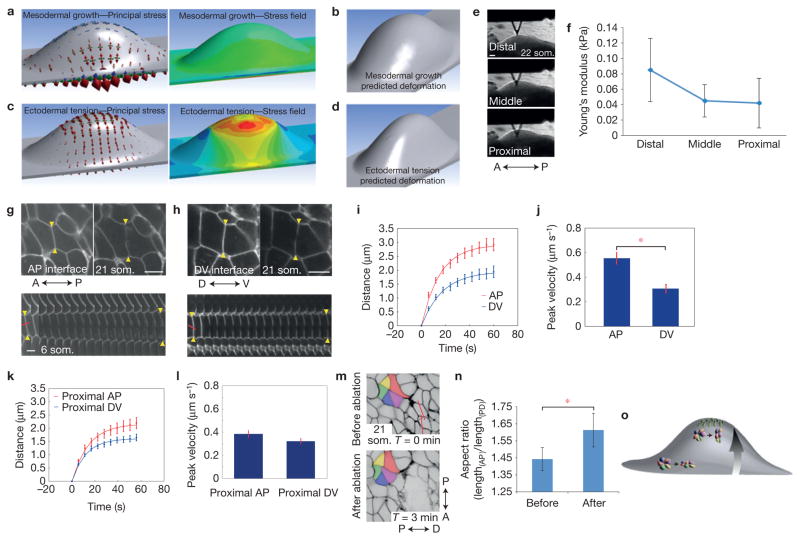
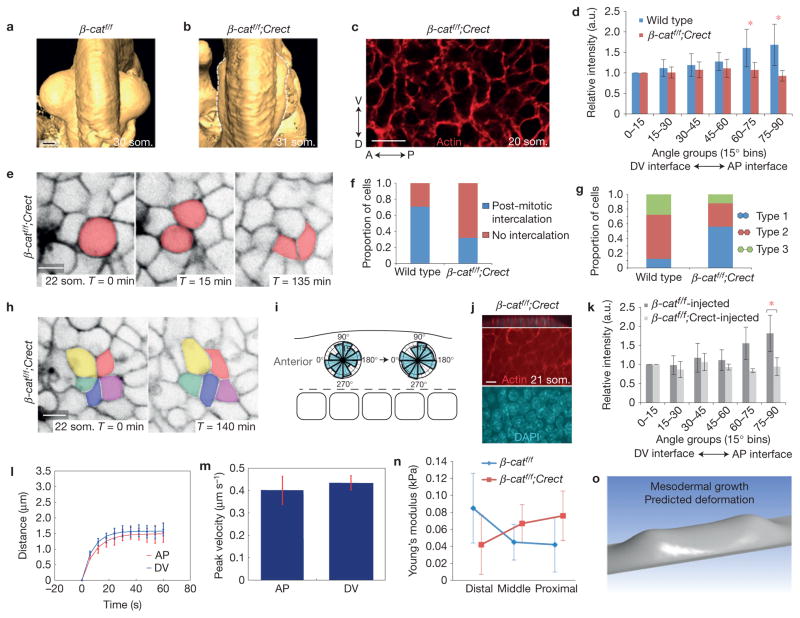
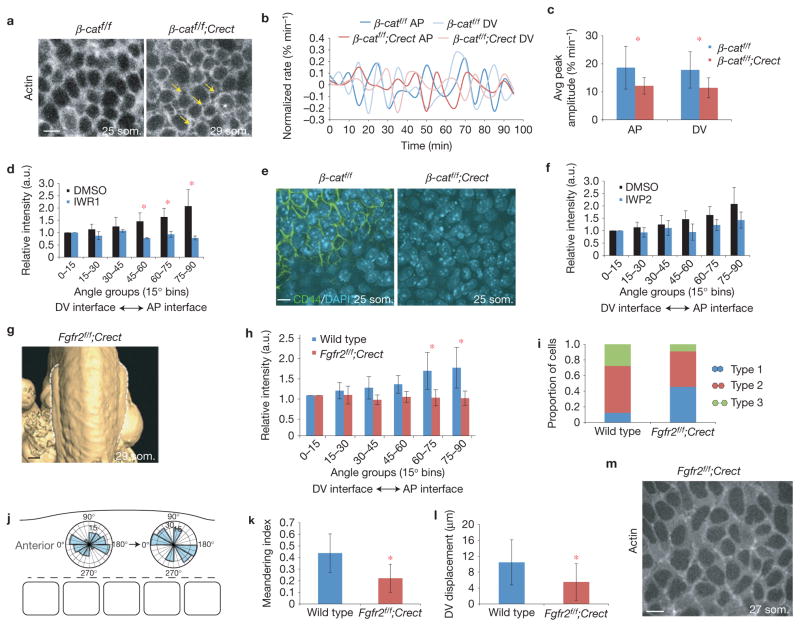
The physical forces that drive morphogenesis are not well characterized in vivo, especially among vertebrates. In the early limb bud, dorsal and ventral ectoderm converge to form the apical ectodermal ridge (AER), although the underlying mechanisms are unclear. By live imaging mouse embryos, we show that prospective AER progenitors intercalate at the dorsoventral boundary and that ectoderm remodels by concomitant cell division and neighbour exchange. Mesodermal expansion and ectodermal tension together generate a dorsoventrally biased stress pattern that orients ectodermal remodelling. Polarized distribution of cortical actin reflects this stress pattern in a β-catenin- and Fgfr2-dependent manner. Intercalation of AER progenitors generates a tensile gradient that reorients resolution of multicellular rosettes on adjacent surfaces, a process facilitated by β-catenin-dependent attachment of cortex to membrane. Therefore, feedback between tissue stress pattern and cell intercalations remodels mammalian ectoderm.
Conflict of interest statement
COMPETING FINANCIAL INTERESTS
The authors declare no competing financial interests.
Figures
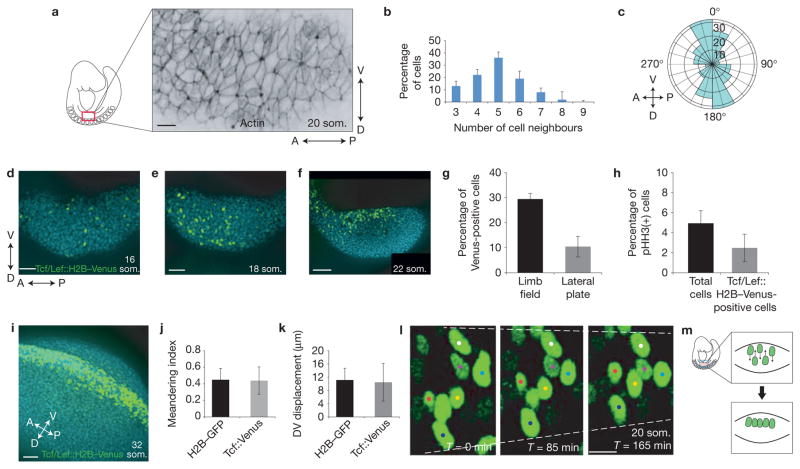
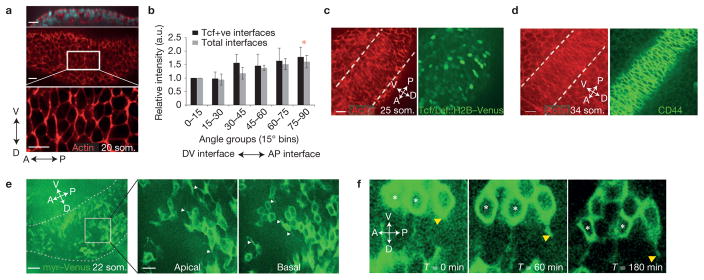
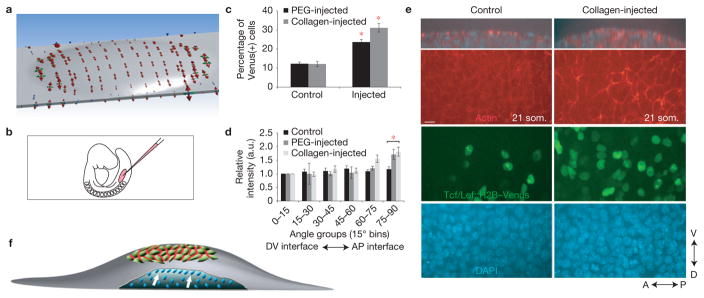
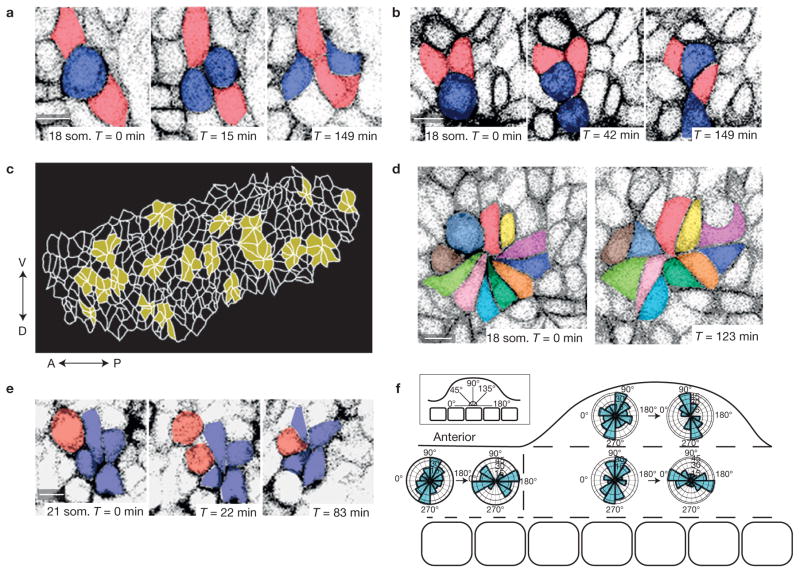
References
-
- Keller R. Developmental biology. Physical biology returns to morphogenesis. Science. 2012;338:201–203. - PubMed
-
- Heisenberg CP, Bellaiche Y. Forces in tissue morphogenesis and patterning. Cell. 2013;153:948–962. - PubMed
-
- Hopyan S, Sharpe J, Yang Y. Budding behaviors: growth of the limb as a model of morphogenesis. Dev Dyn. 2011;240:1054–1062. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
