

An Invertron-Like Linear Plasmid Mediates Intracellular Survival and Virulence in Bovine Isolates of Rhodococcus equi
- PMID: 25895973
- PMCID: PMC4468562
- DOI: 10.1128/IAI.00376-15
An Invertron-Like Linear Plasmid Mediates Intracellular Survival and Virulence in Bovine Isolates of Rhodococcus equi
Abstract
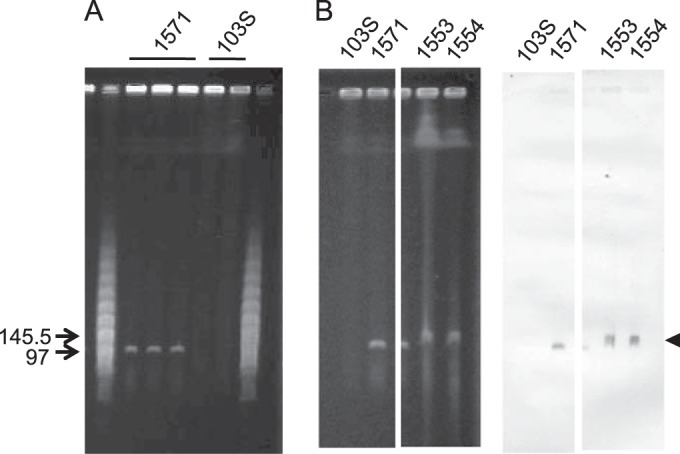
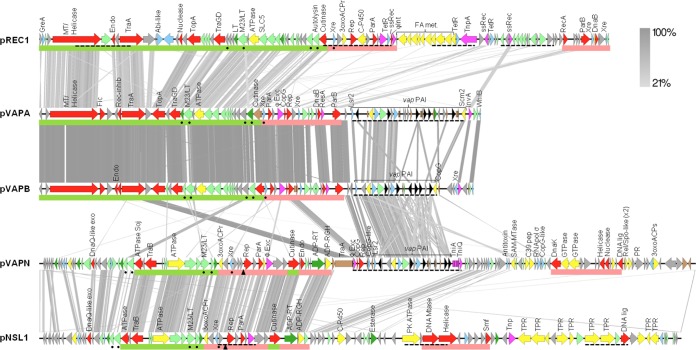
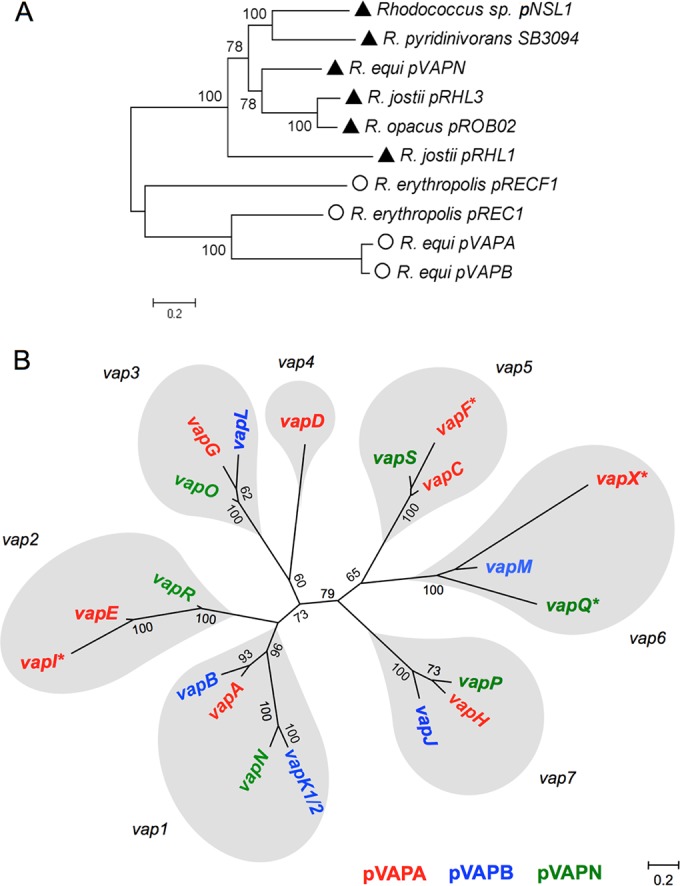
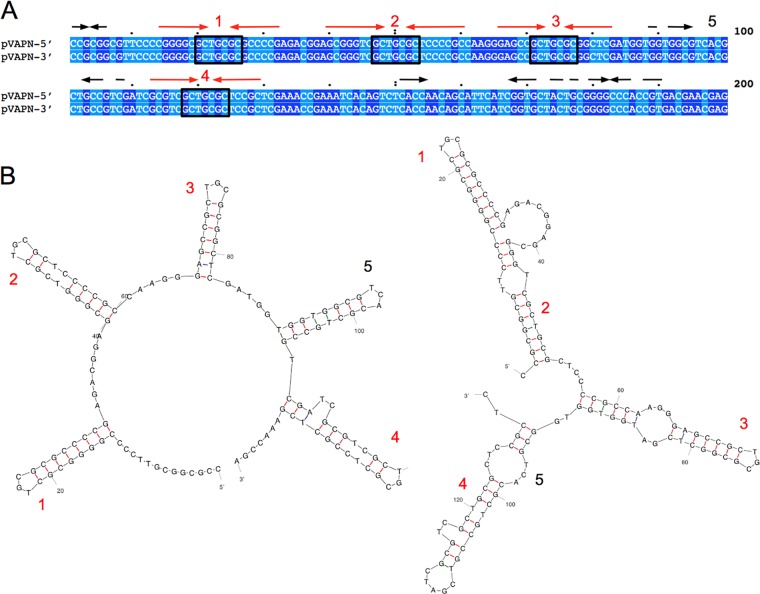
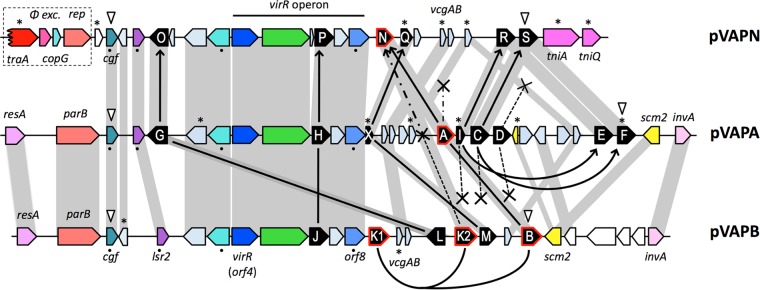
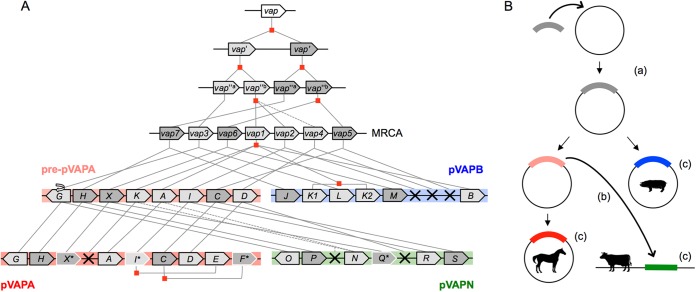
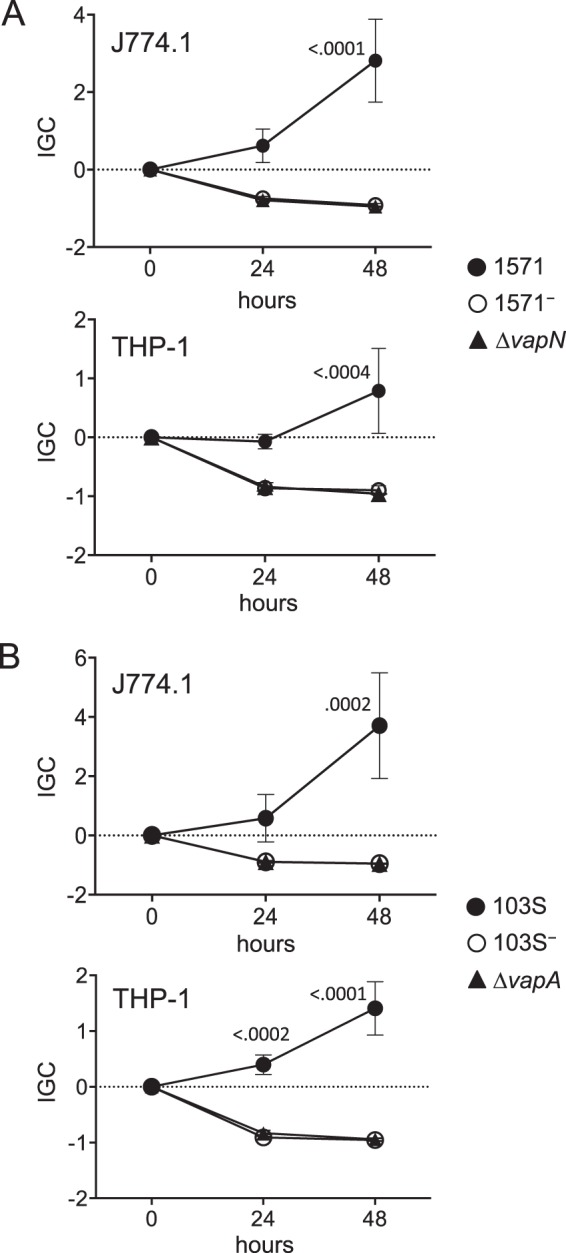
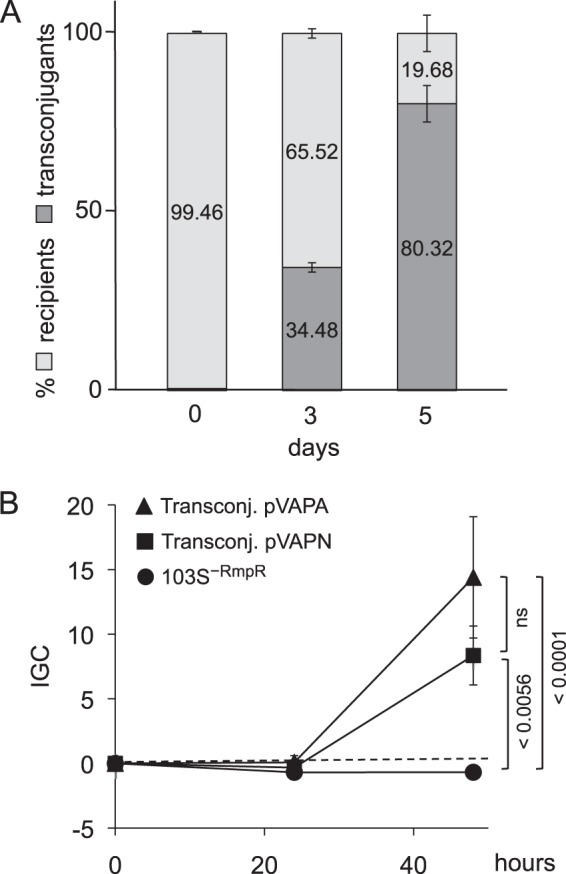
We report a novel host-associated virulence plasmid in Rhodococcus equi, pVAPN, carried by bovine isolates of this facultative intracellular pathogenic actinomycete. Surprisingly, pVAPN is a 120-kb invertron-like linear replicon unrelated to the circular virulence plasmids associated with equine (pVAPA) and porcine (pVAPB variant) R. equi isolates. pVAPN is similar to the linear plasmid pNSL1 from Rhodococcus sp. NS1 and harbors six new vap multigene family members (vapN to vapS) in a vap pathogenicity locus presumably acquired via en bloc mobilization from a direct predecessor of equine pVAPA. Loss of pVAPN rendered R. equi avirulent in macrophages and mice. Mating experiments using an in vivo transconjugant selection strategy demonstrated that pVAPN transfer is sufficient to confer virulence to a plasmid-cured R. equi recipient. Phylogenetic analyses assigned the vap multigene family complement from pVAPN, pVAPA, and pVAPB to seven monophyletic clades, each containing plasmid type-specific allelic variants of a precursor vap gene carried by the nearest vap island ancestor. Deletion of vapN, the predicted "bovine-type" allelic counterpart of vapA, essential for virulence in pVAPA, abrogated pVAPN-mediated intramacrophage proliferation and virulence in mice. Our findings support a model in which R. equi virulence is conferred by host-adapted plasmids. Their central role is mediating intracellular proliferation in macrophages, promoted by a key vap determinant present in the common ancestor of the plasmid-specific vap islands, with host tropism as a secondary trait selected during coevolution with specific animal species.
Copyright © 2015, American Society for Microbiology. All Rights Reserved.
Figures

References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
