

Involvement of unconventional myosin VI in myoblast function and myotube formation
- PMID: 25896210
- PMCID: PMC4469105
- DOI: 10.1007/s00418-015-1322-6
Involvement of unconventional myosin VI in myoblast function and myotube formation
Abstract
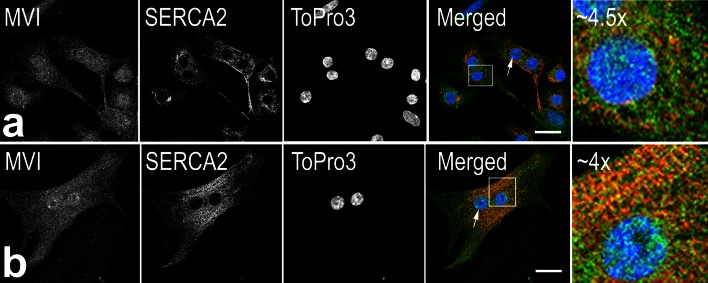
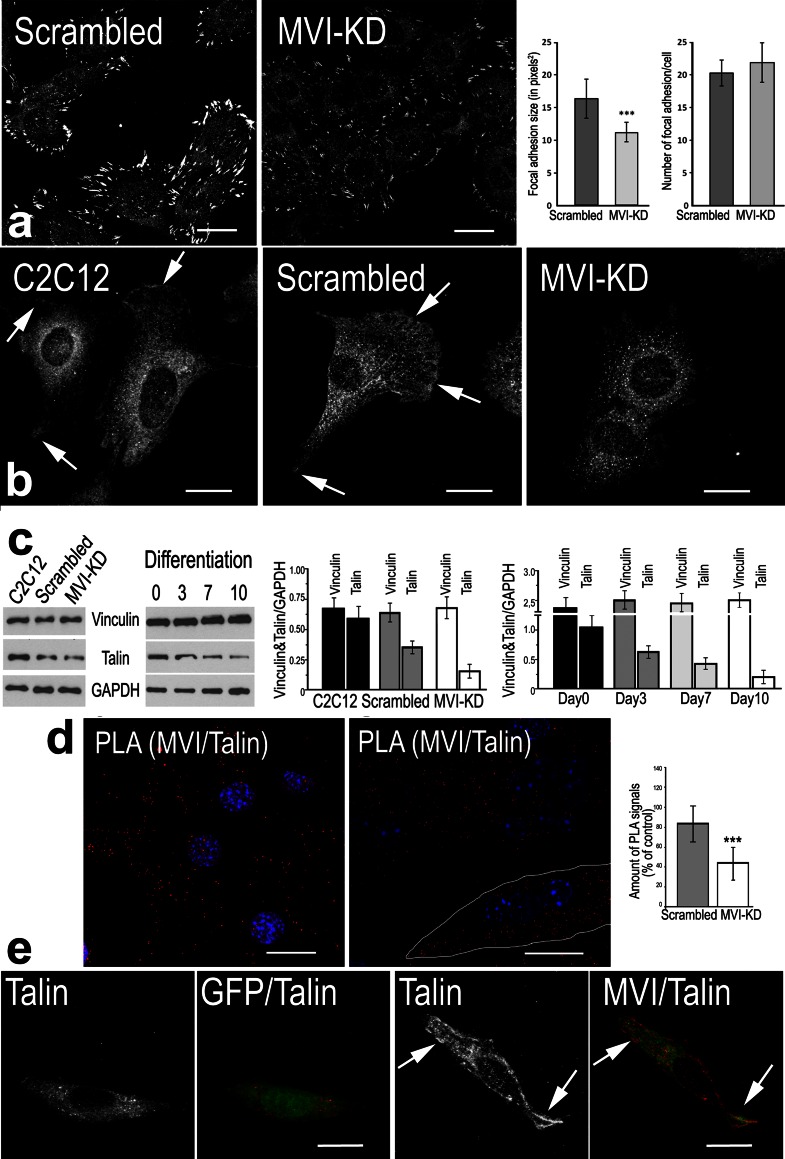
The important role of unconventional myosin VI (MVI) in skeletal and cardiac muscle has been recently postulated (Karolczak et al. in Histochem Cell Biol 139:873-885, 2013). Here, we addressed for the first time a role for this unique myosin motor in myogenic cells as well as during their differentiation into myotubes. During myoblast differentiation, the isoform expression pattern of MVI and its subcellular localization underwent changes. In undifferentiated myoblasts, MVI-stained puncti were seen throughout the cytoplasm and were in close proximity to actin filaments, Golgi apparatus, vinculin-, and talin-rich focal adhesion as well as endoplasmic reticulum. Colocalization of MVI with endoplasmic reticulum was enhanced during myotube formation, and differentiation-dependent association was also seen in sarcoplasmic reticulum of neonatal rat cardiomyocytes (NRCs). Moreover, we observed enrichment of MVI in myotube regions containing acetylcholine receptor-rich clusters, suggesting its involvement in the organization of the muscle postsynaptic machinery. Overexpression of the H246R MVI mutant (associated with hypertrophic cardiomyopathy) in myoblasts and NRCs caused the formation of abnormally large intracellular vesicles. MVI knockdown caused changes in myoblast morphology and inhibition of their migration. On the subcellular level, MVI-depleted myoblasts exhibited aberrations in the organization of actin cytoskeleton and adhesive structures as well as in integrity of Golgi apparatus and endoplasmic reticulum. Also, MVI depletion or overexpression of H246R mutant caused the formation of significantly wider or aberrant myotubes, respectively, indicative of involvement of MVI in myoblast differentiation. The presented results suggest an important role for MVI in myogenic cells and possibly in myoblast differentiation.
Figures








References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
