

Visual representations are dominated by intrinsic fluctuations correlated between areas
- PMID: 25896934
- PMCID: PMC4503804
- DOI: 10.1016/j.neuroimage.2015.04.026
Visual representations are dominated by intrinsic fluctuations correlated between areas
Abstract
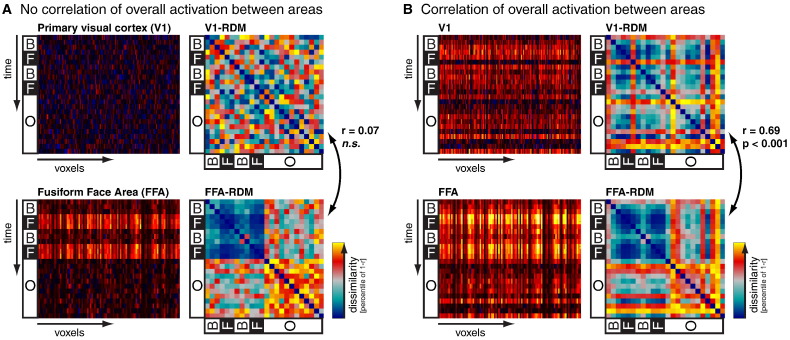
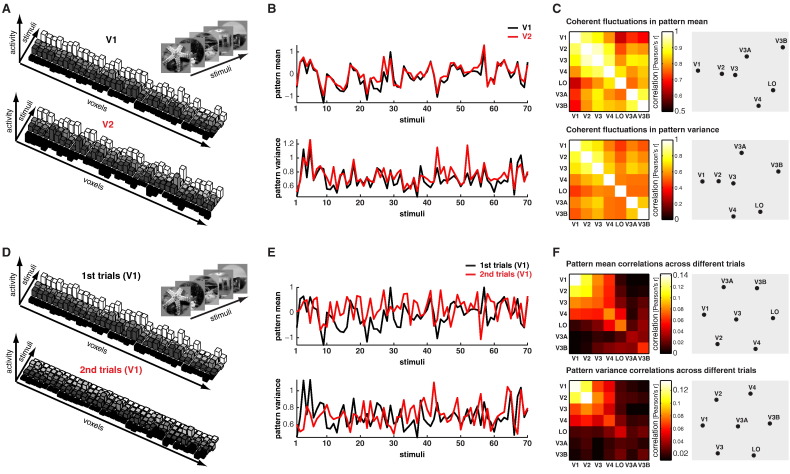
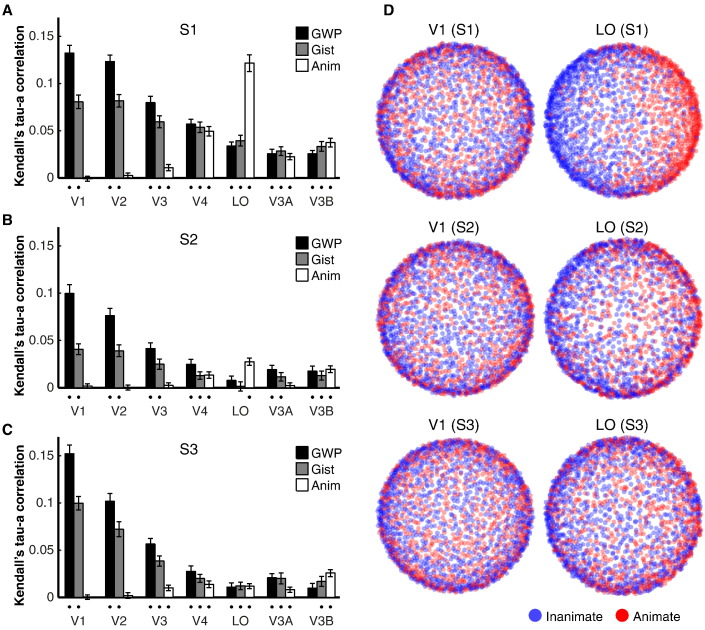
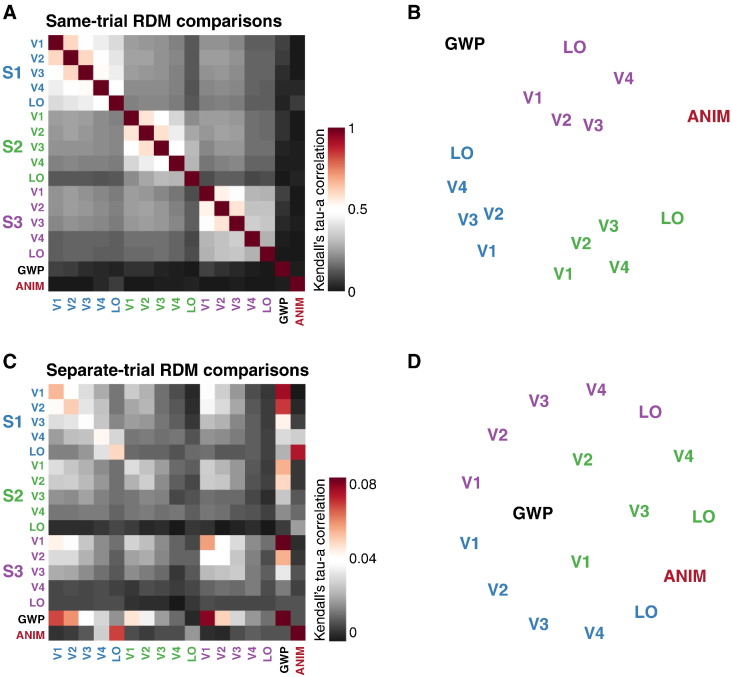
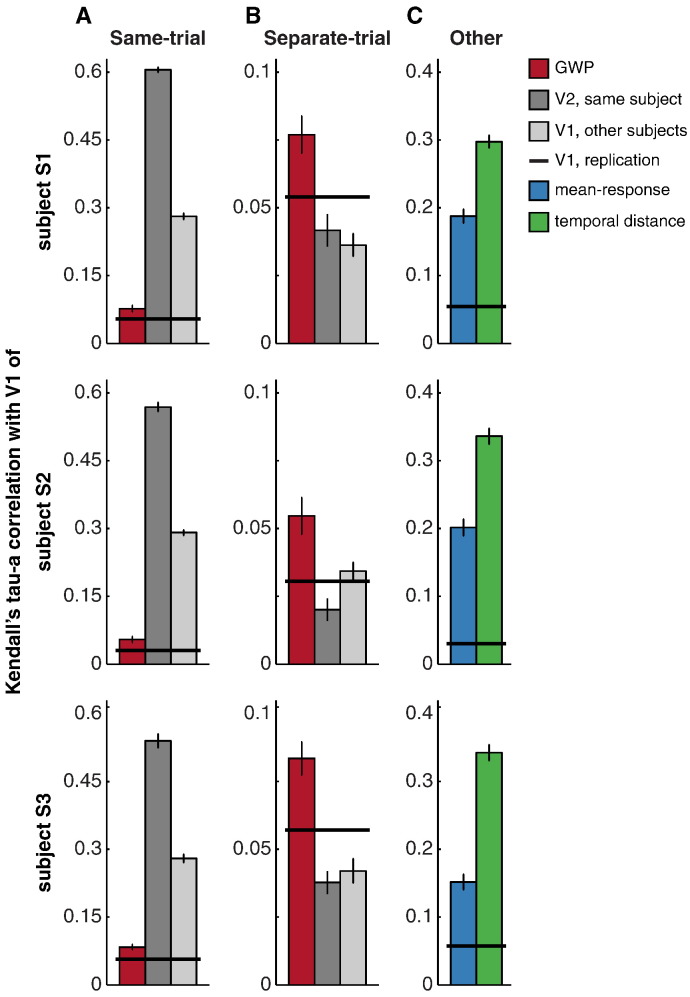
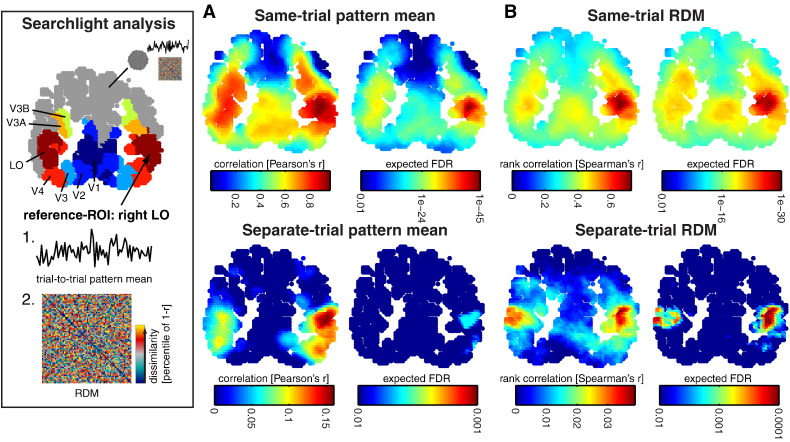
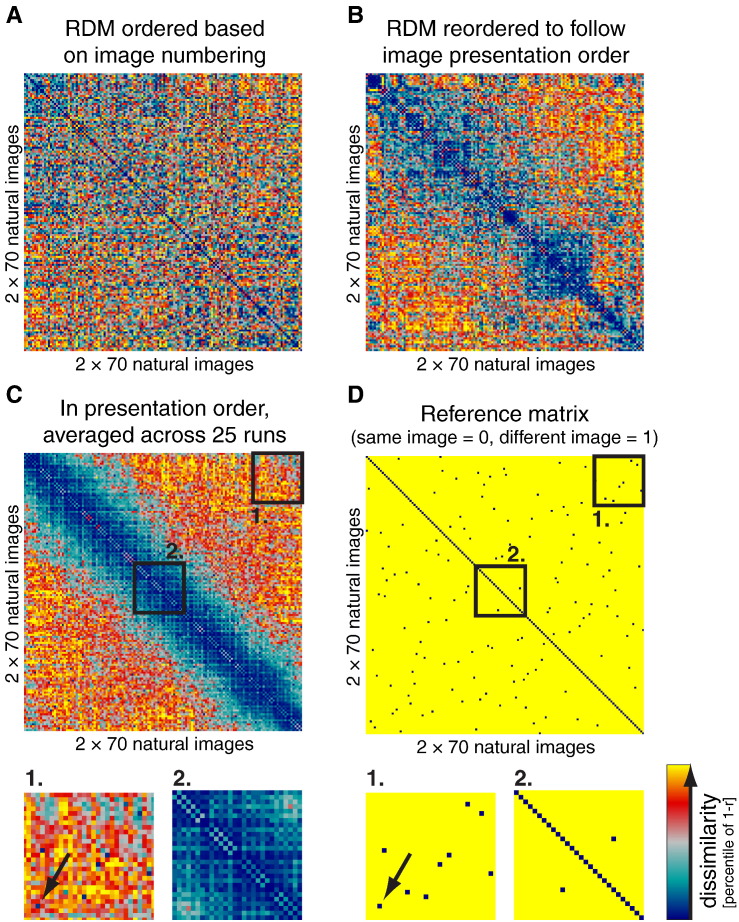
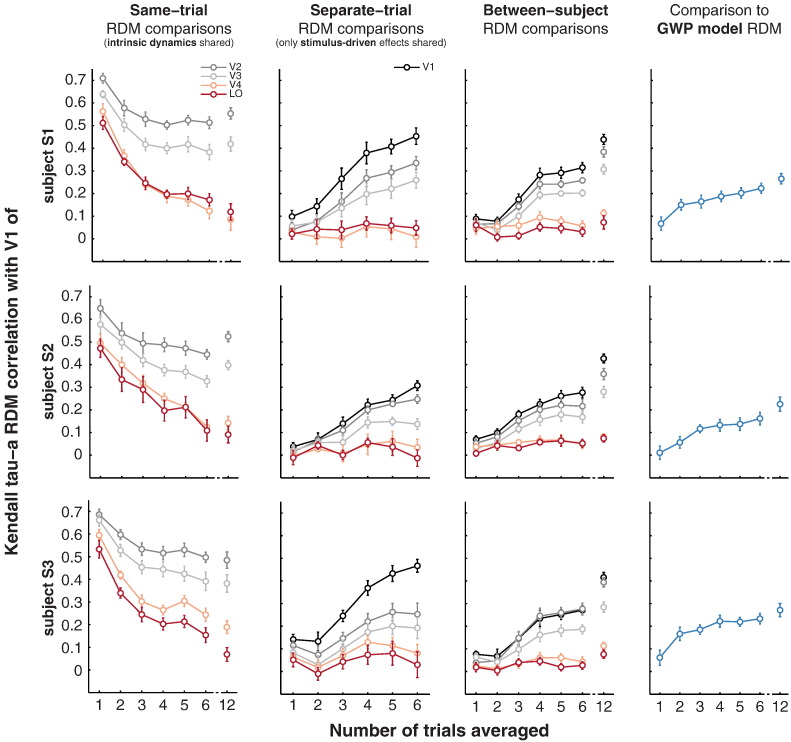
Intrinsic cortical dynamics are thought to underlie trial-to-trial variability of visually evoked responses in animal models. Understanding their function in the context of sensory processing and representation is a major current challenge. Here we report that intrinsic cortical dynamics strongly affect the representational geometry of a brain region, as reflected in response-pattern dissimilarities, and exaggerate the similarity of representations between brain regions. We characterized the representations in several human visual areas by representational dissimilarity matrices (RDMs) constructed from fMRI response-patterns for natural image stimuli. The RDMs of different visual areas were highly similar when the response-patterns were estimated on the basis of the same trials (sharing intrinsic cortical dynamics), and quite distinct when patterns were estimated on the basis of separate trials (sharing only the stimulus-driven component). We show that the greater similarity of the representational geometries can be explained by coherent fluctuations of regional-mean activation within visual cortex, reflecting intrinsic dynamics. Using separate trials to study stimulus-driven representations revealed clearer distinctions between the representational geometries: a Gabor wavelet pyramid model explained representational geometry in visual areas V1-3 and a categorical animate-inanimate model in the object-responsive lateral occipital cortex.
Keywords: Functional MRI; Intrinsic dynamics; Natural images; Pattern information; Representational similarity; Visual cortex.
Copyright © 2015. Published by Elsevier Inc.
Figures


References
-
- Arieli A., Sterkin A., Grinvald A., Aertsen A. Dynamics of ongoing activity: explanation of the large variability in evoked cortical responses. Science. 1996;273:1868–1871. - PubMed
-
- Cavanaugh J.R., Bair W., Movshon J.A. Nature and interaction of signals from the receptive field center and surround in macaque V1 neurons. J. Neurophysiol. 2002;88:2530–2546. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
