

The Association of the Xeroderma Pigmentosum Group D DNA Helicase (XPD) with Transcription Factor IIH Is Regulated by the Cytosolic Iron-Sulfur Cluster Assembly Pathway
- PMID: 25897079
- PMCID: PMC4447990
- DOI: 10.1074/jbc.M115.650762
The Association of the Xeroderma Pigmentosum Group D DNA Helicase (XPD) with Transcription Factor IIH Is Regulated by the Cytosolic Iron-Sulfur Cluster Assembly Pathway
Abstract
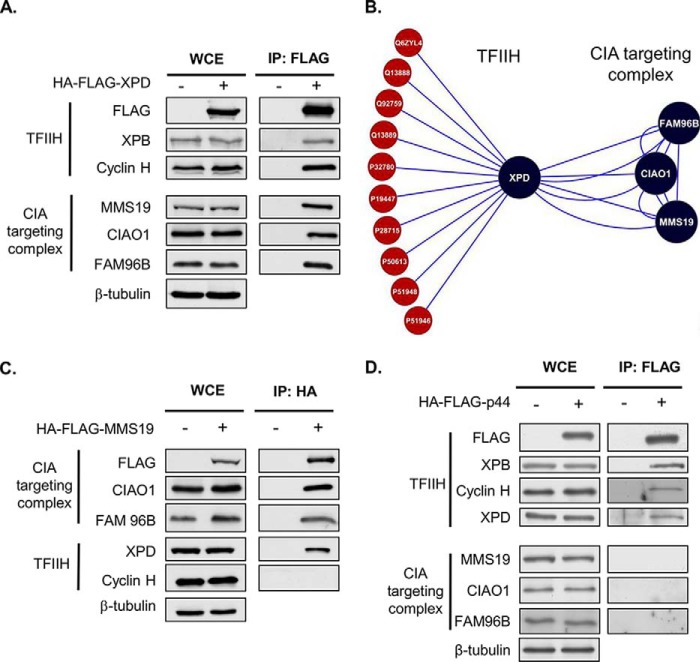
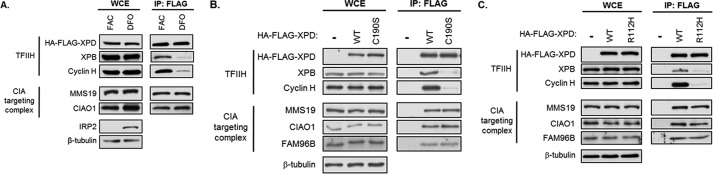
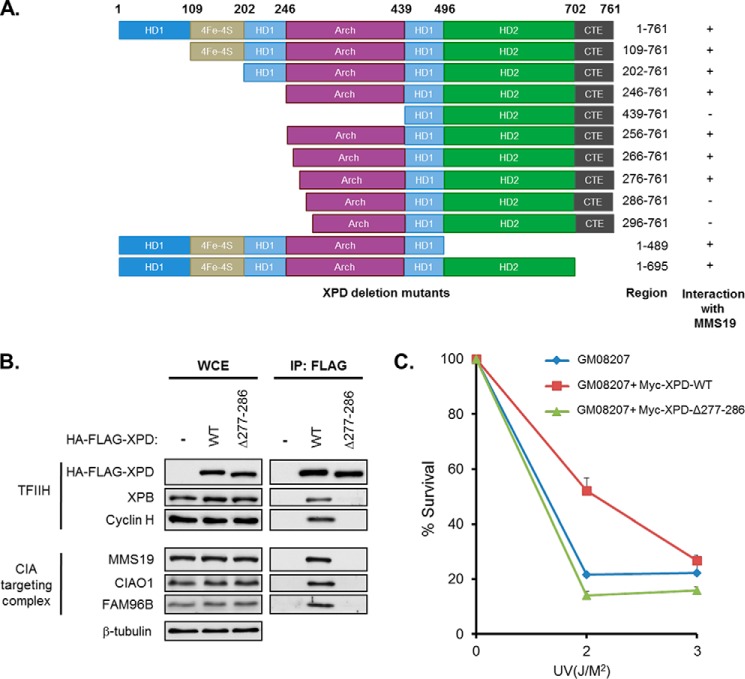
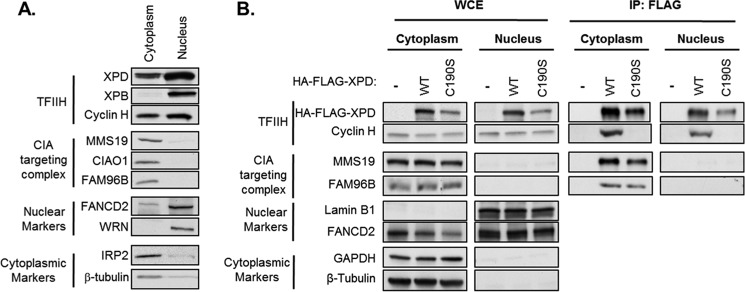
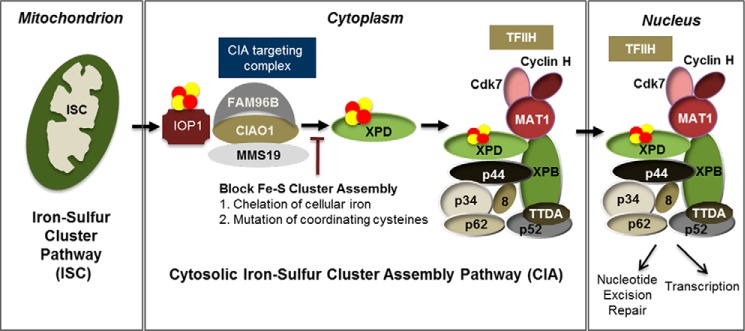
Xeroderma pigmentosum group D (XPD) helicase is a component of the transcription factor IIH (TFIIH) transcription complex and plays essential roles in transcription and nucleotide excision repair. Although iron-sulfur (Fe-S) cluster binding by XPD is required for activity, the process mediating Fe-S cluster assembly remains poorly understood. We recently identified a cytoplasmic Fe-S cluster assembly (CIA) targeting complex composed of MMS19, CIAO1, and FAM96B that is required for the biogenesis of extramitochondrial Fe-S proteins including XPD. Here, we use XPD as a prototypical Fe-S protein to further characterize how Fe-S assembly is facilitated by the CIA targeting complex. Multiple lines of evidence indicate that this process occurs in a stepwise fashion in which XPD acquires a Fe-S cluster from the CIA targeting complex before assembling into TFIIH. First, XPD was found to associate in a mutually exclusive fashion with either TFIIH or the CIA targeting complex. Second, disrupting Fe-S cluster assembly on XPD by either 1) depleting cellular iron levels or 2) utilizing XPD mutants defective in either Fe-S cluster or CIA targeting complex binding blocks Fe-S cluster assembly and prevents XPD incorporation into TFIIH. Finally, subcellular fractionation studies indicate that the association of XPD with the CIA targeting complex occurs in the cytoplasm, whereas its association with TFIIH occurs largely in the nucleus where TFIIH functions. Together, these data establish a sequential assembly process for Fe-S assembly on XPD and highlight the existence of quality control mechanisms that prevent the incorporation of immature apoproteins into their cellular complexes.
Keywords: DNA helicase; DNA repair; iron metabolism; iron-sulfur protein; proteomics.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Friedberg E. C. (2001) How nucleotide excision repair protects against cancer. Nat. Rev. Cancer 1, 22–33 - PubMed
-
- Lehmann A. R. (2001) The xeroderma pigmentosum group D (XPD) gene: one gene, two functions, three diseases. Genes Dev. 15, 15–23 - PubMed
-
- Netz D. J., Mascarenhas J., Stehling O., Pierik A. J., Lill R. (2014) Maturation of cytosolic and nuclear iron-sulfur proteins. Trends Cell Biol. 24, 303–312 - PubMed
-
- Lill R. (2009) Function and biogenesis of iron-sulphur proteins. Nature 460, 831–838 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
