

Collagenolytic Matrix Metalloproteinase Activities toward Peptomeric Triple-Helical Substrates
- PMID: 25897652
- PMCID: PMC4511165
- DOI: 10.1021/acs.biochem.5b00110
Collagenolytic Matrix Metalloproteinase Activities toward Peptomeric Triple-Helical Substrates
Abstract
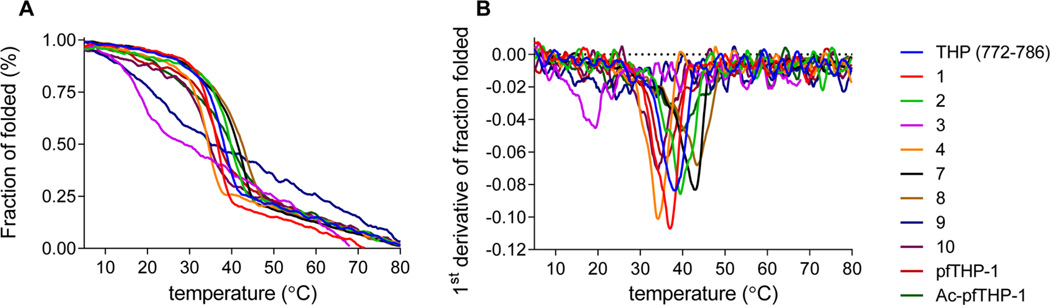
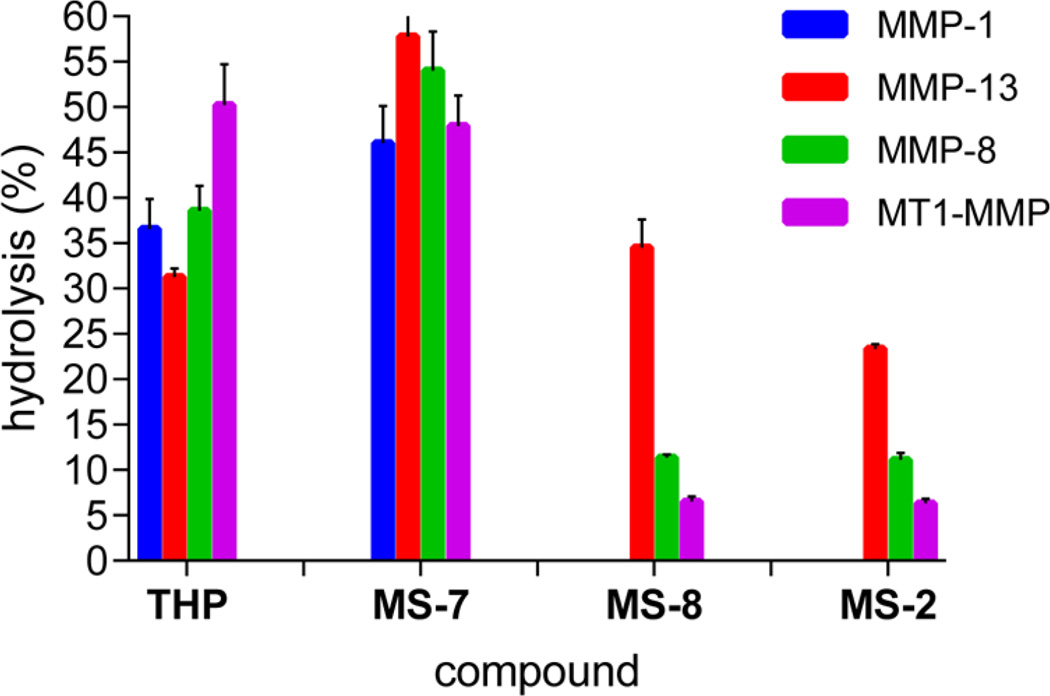
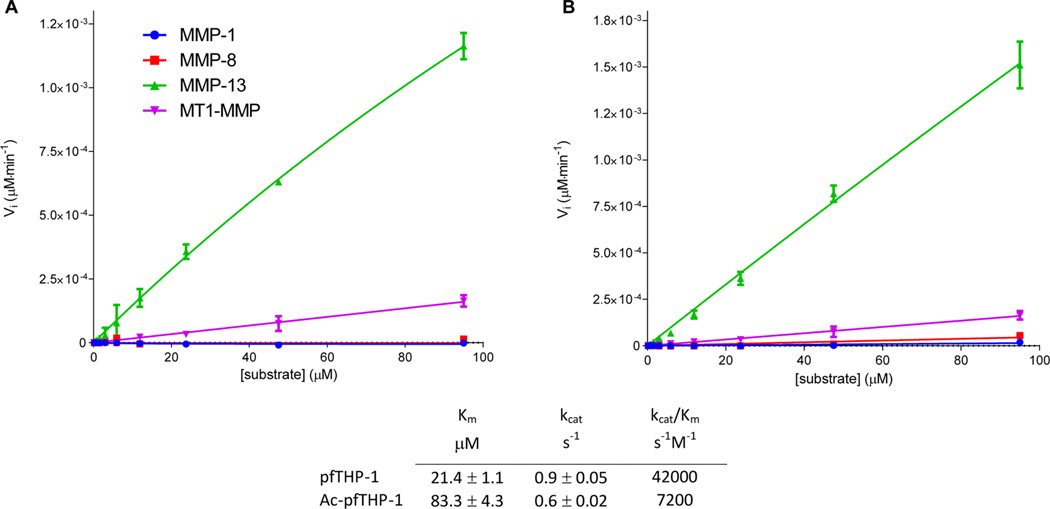
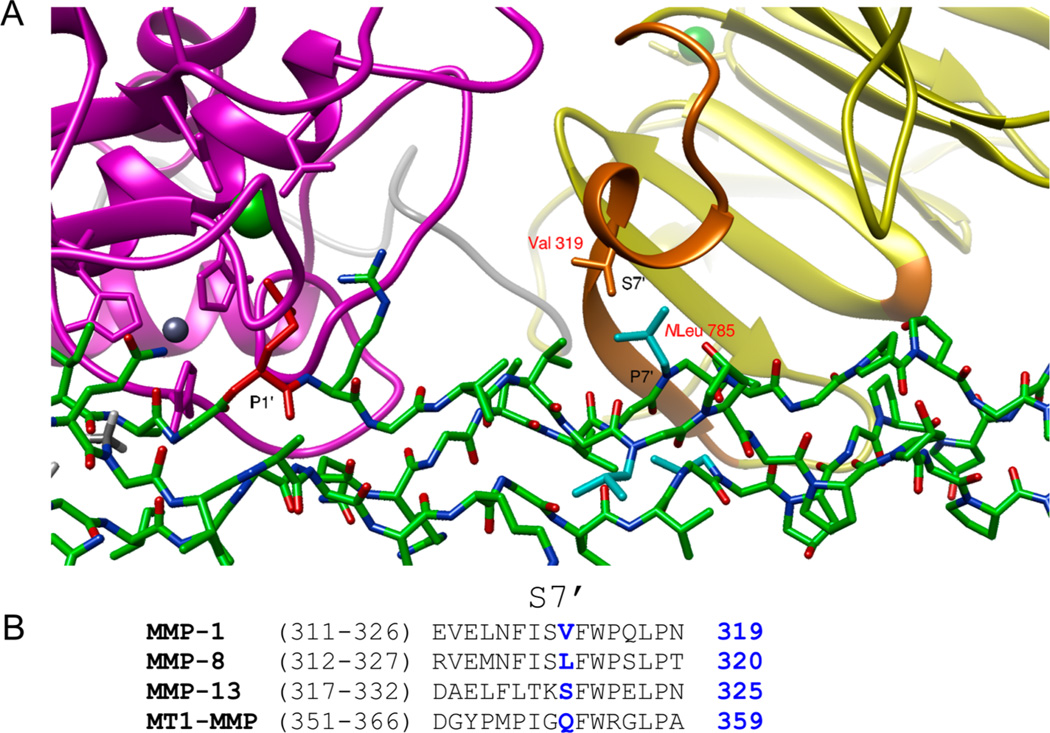
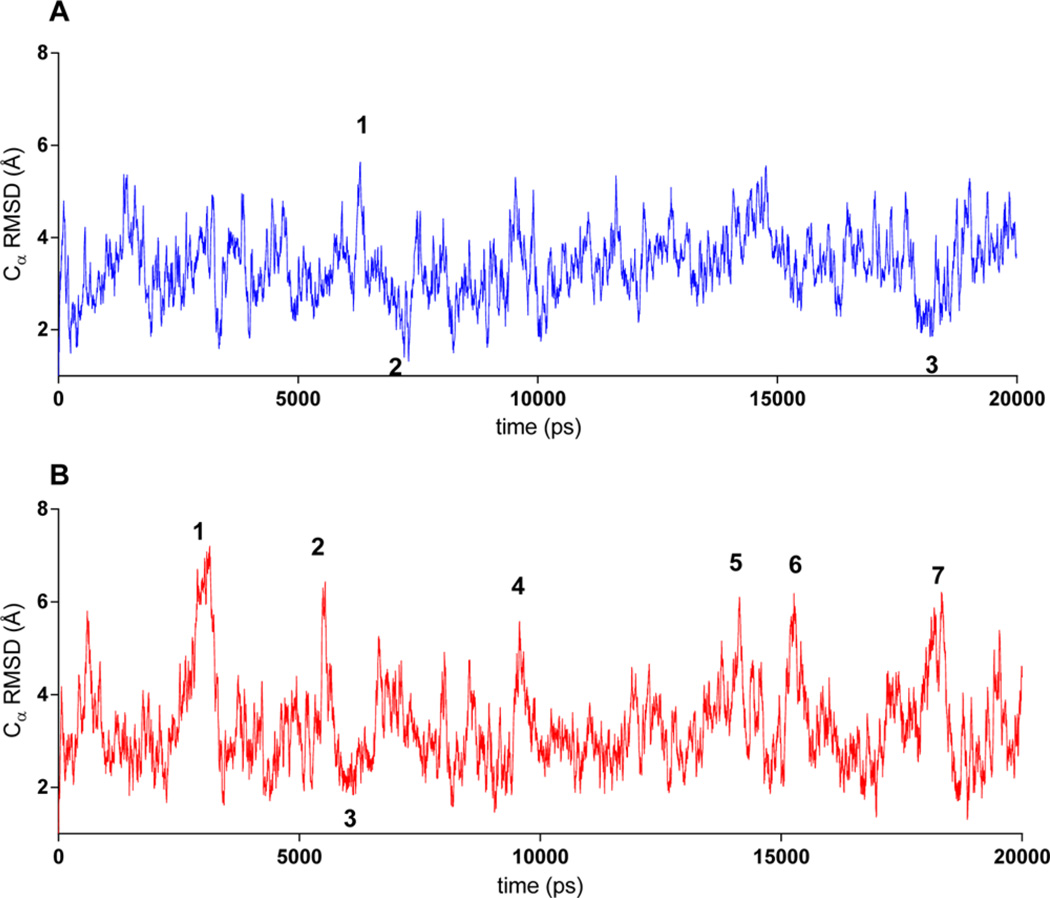
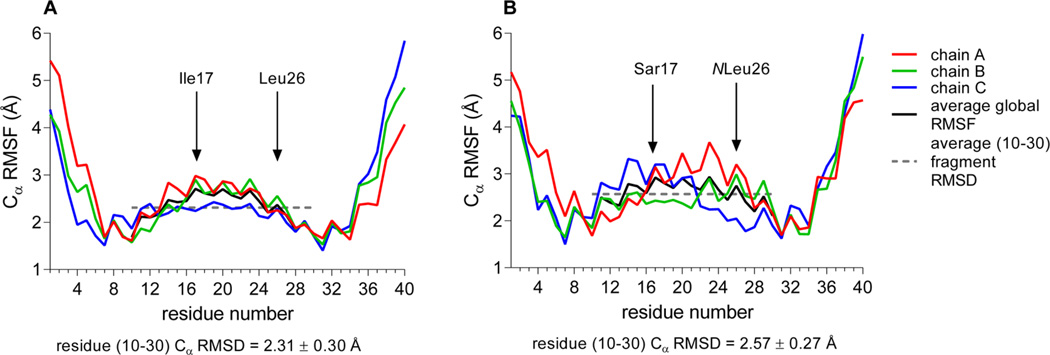
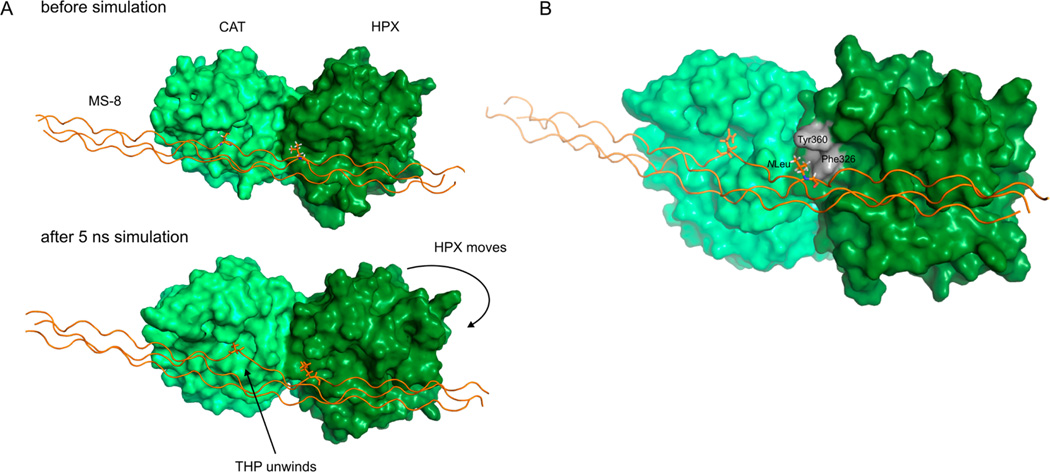
Although collagenolytic matrix metalloproteinases (MMPs) possess common domain organizations, there are subtle differences in their processing of collagenous triple-helical substrates. In this study, we have incorporated peptoid residues into collagen model triple-helical peptides and examined MMP activities toward these peptomeric chimeras. Several different peptoid residues were incorporated into triple-helical substrates at subsites P3, P1, P1', and P10' individually or in combination, and the effects of the peptoid residues were evaluated on the activities of full-length MMP-1, MMP-8, MMP-13, and MMP-14/MT1-MMP. Most peptomers showed little discrimination between MMPs. However, a peptomer containing N-methyl Gly (sarcosine) in the P1' subsite and N-isobutyl Gly (NLeu) in the P10' subsite was hydrolyzed efficiently only by MMP-13 [nomenclature relative to the α1(I)772-786 sequence]. Cleavage site analysis showed hydrolysis at the Gly-Gln bond, indicating a shifted binding of the triple helix compared to the parent sequence. Favorable hydrolysis by MMP-13 was not due to sequence specificity or instability of the substrate triple helix but rather was based on the specific interactions of the P7' peptoid residue with the MMP-13 hemopexin-like domain. A fluorescence resonance energy transfer triple-helical peptomer was constructed and found to be readily processed by MMP-13, not cleaved by MMP-1 and MMP-8, and weakly hydrolyzed by MT1-MMP. The influence of the triple-helical structure containing peptoid residues on the interaction between MMP subsites and individual substrate residues may provide additional information about the mechanism of collagenolysis, the understanding of collagen specificity, and the design of selective MMP probes.
Figures


References
-
- McCawley LJ, Matrisian LM. Matrix metalloproteinases: They’re not just for matrix anymore. Curr. Opin. Cell Biol. 2001;13:534–540. - PubMed
-
- Overall CM. Molecular determinants of metalloproteinase substrate specificity. Mol. Biotechnol. 2002;22:51–86. - PubMed
-
- Morrison CJ, Overall CM. TIMP-independence of MMP-2 activation by MT2-MMP is determined by contributions of both the MT2-MMP catalytic and hemopexin domains. J. Biol. Chem. 2006;281:26528–26539. - PubMed
-
- Bigg HF, Rowan AD, Barker MD, Cawston TE. Activity of matrix metalloproteinase-9 against native collagen types I and III. FEBS J. 2007;274:1246–1255. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
