

Manganese homeostasis and utilization in pathogenic bacteria
- PMID: 25898914
- PMCID: PMC4631260
- DOI: 10.1111/mmi.13034
Manganese homeostasis and utilization in pathogenic bacteria
Abstract
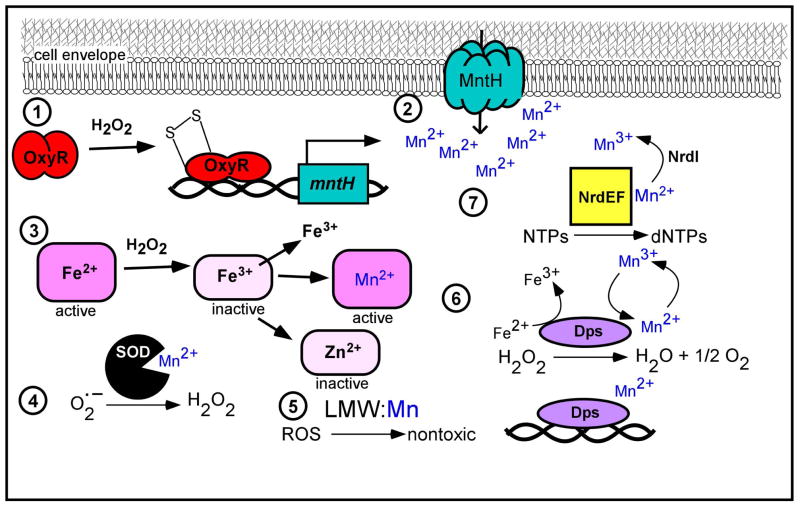
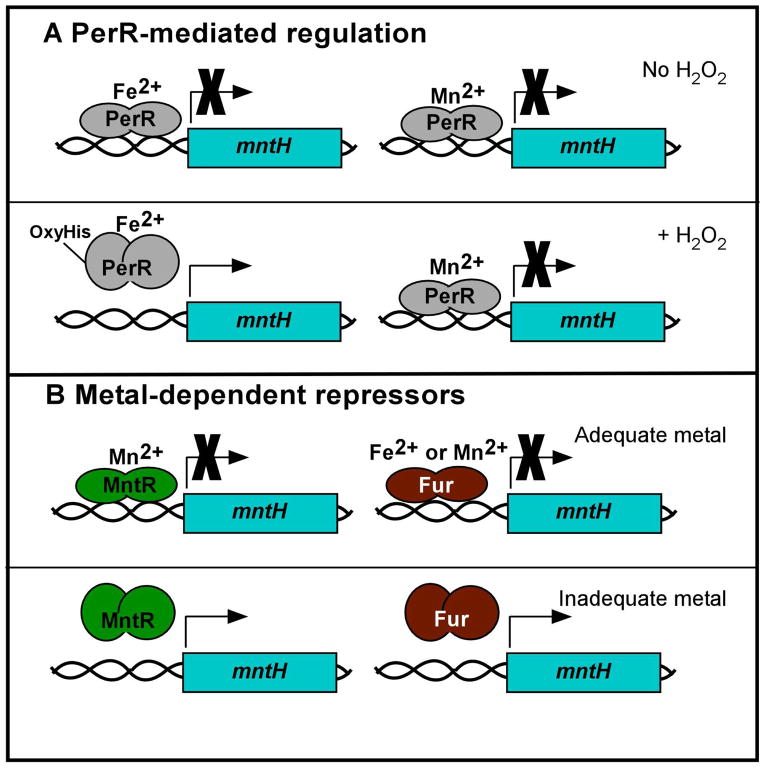
Manganese (Mn) is a required cofactor for all forms of life. Given the importance of Mn to bacteria, the host has devised strategies to sequester Mn from invaders. In the macrophage phagosome, NRAMP1 removes Mn and other essential metals to starve intracellular pathogens; in the extracellular space, calprotectin chelates Mn and Zn. Calprotectin-mediated Mn sequestration is a newly appreciated host defense mechanism, and recent findings are highlighted herein. In order to acquire Mn when extracellular concentrations are low, bacteria have evolved efficient Mn acquisition systems that are under elegant transcriptional control. To counteract Mn overload, some bacteria possess Mn-specific export systems that are important in vivo, presumably for control of intracellular Mn levels. Mn transporters, their transcriptional regulators and some Mn-requiring enzymes are necessary for virulence of certain bacterial pathogens, as revealed by animal models of infection. Furthermore, Mn is an important facet of the cellular response to oxidative stress, a host antibacterial strategy. The battle for Mn between host and pathogen is now appreciated to be a major determinant of the outcome of infection. In this MicroReview, the contribution of Mn to the host-pathogen interaction is reviewed, and key questions are proposed for future study.
© 2015 John Wiley & Sons Ltd.
Figures

References
-
- Achouiti A, Vogl T, Endeman H, Mortensen BL, Laterre PF, Wittebole X, et al. Myeloid-related protein-8/14 facilitates bacterial growth during pneumococcal pneumonia. Thorax. 2014;69:1034–1042. - PubMed
-
- Almiron M, Link AJ, Furlong D, Kolter R. A novel DNA-binding protein with regulatory and protective roles in starved Escherichia coli. Genes Dev. 1992;6:2646–2654. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
