

The Psp system of Mycobacterium tuberculosis integrates envelope stress-sensing and envelope-preserving functions
- PMID: 25899163
- PMCID: PMC4642893
- DOI: 10.1111/mmi.13037
The Psp system of Mycobacterium tuberculosis integrates envelope stress-sensing and envelope-preserving functions
Abstract
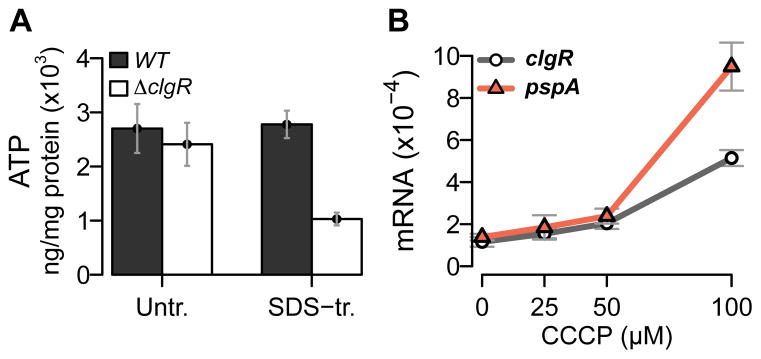
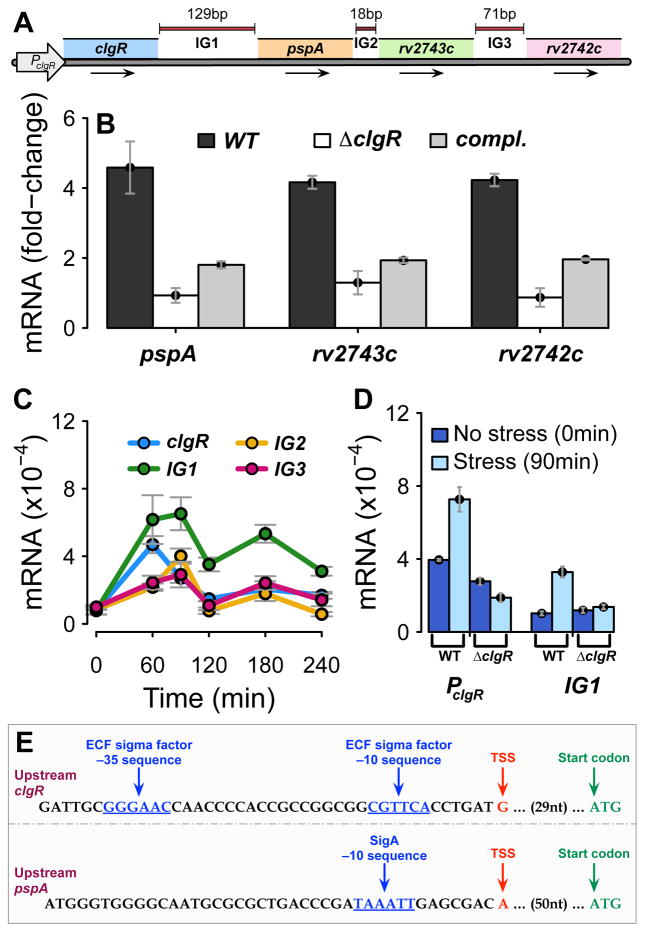
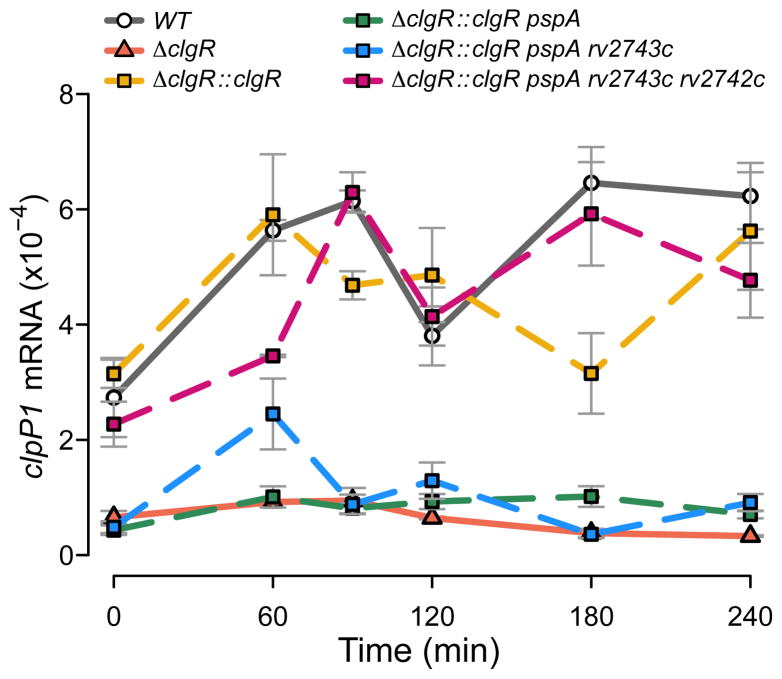
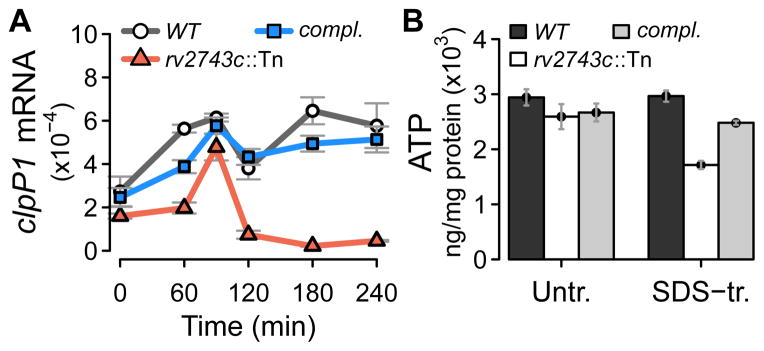
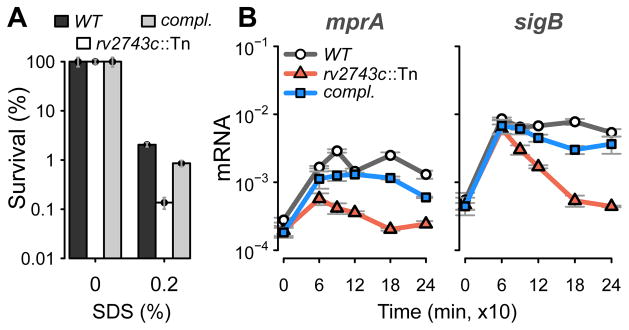
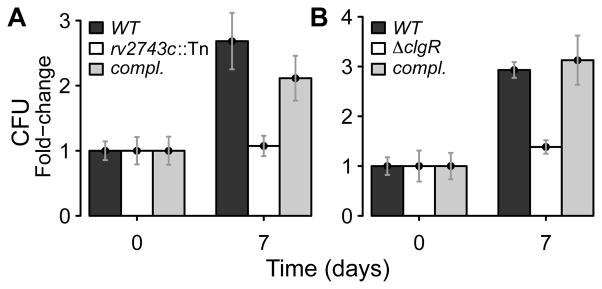
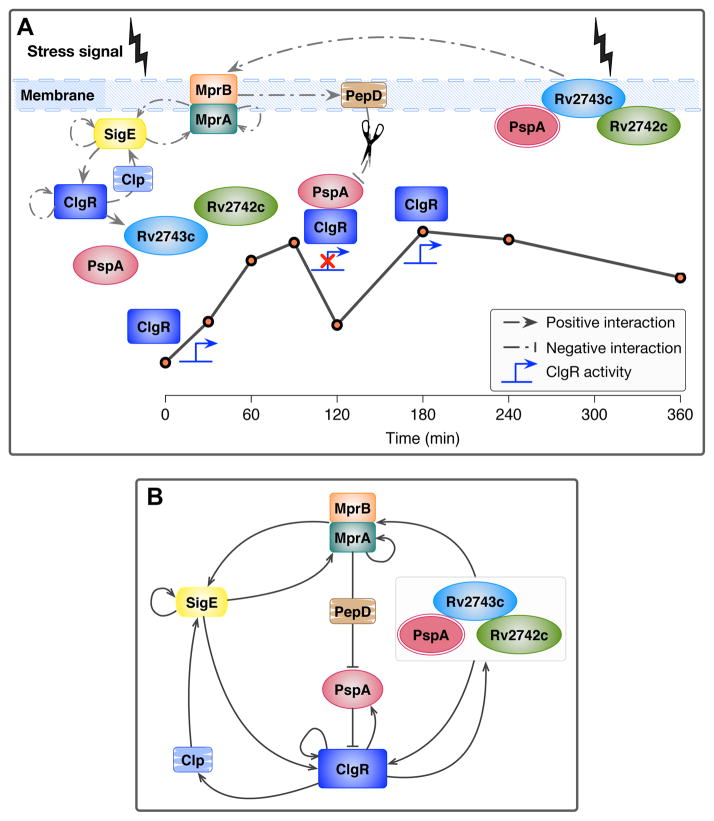
The bacterial envelope integrates essential stress-sensing and adaptive functions; thus, envelope-preserving functions are important for survival. In Gram-negative bacteria, envelope integrity during stress is maintained by the multi-gene Psp response. Mycobacterium tuberculosis was thought to lack the Psp system since it encodes only pspA and no other psp ortholog. Intriguingly, pspA maps downstream from clgR, which encodes a transcription factor regulated by the MprAB-σ(E) envelope-stress-signaling system. clgR inactivation lowered ATP concentration during stress and protonophore treatment-induced clgR-pspA expression, suggesting that these genes express Psp-like functions. We identified a four-gene set - clgR, pspA (rv2744c), rv2743c, rv2742c - that is regulated by clgR and in turn regulates ClgR activity. Regulatory and protein-protein interactions within the set and a requirement of the four genes for functions associated with envelope integrity and surface-stress tolerance indicate that a Psp-like system has evolved in mycobacteria. Among Actinobacteria, the four-gene module occurred only in tuberculous mycobacteria and was required for intramacrophage growth, suggesting links between its function and mycobacterial virulence. Additionally, the four-gene module was required for MprAB-σ(E) stress-signaling activity. The positive feedback between envelope-stress-sensing and envelope-preserving functions allows sustained responses to multiple, envelope-perturbing signals during chronic infection, making the system uniquely suited to tuberculosis pathogenesis.
© 2015 John Wiley & Sons Ltd.
Conflict of interest statement
The authors have no conflict of interest to declare.
Figures



References
-
- Altschul SF, Gish W, Miller W, Myers EW, Lipman DJ. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Barik S, Sureka K, Mukherjee P, Basu J, Kundu M. RseA, the SigE specific anti-sigma factor of Mycobacterium tuberculosis, is inactivated by phosphorylation-dependent ClpC1P2 proteolysis. Mol Microbiol. 2010;75:592–606. - PubMed
-
- Becker LA, Bang IS, Crouch ML, Fang FC. Compensatory role of PspA, a member of the phage shock protein operon, in rpoE mutant Salmonella enterica serovar Typhimurium. Mol Microbiol. 2005;56:1004–1016. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
