

Diverse involvement of EZH2 in cancer epigenetics
- PMID: 25901190
- PMCID: PMC4399085
Diverse involvement of EZH2 in cancer epigenetics
Abstract
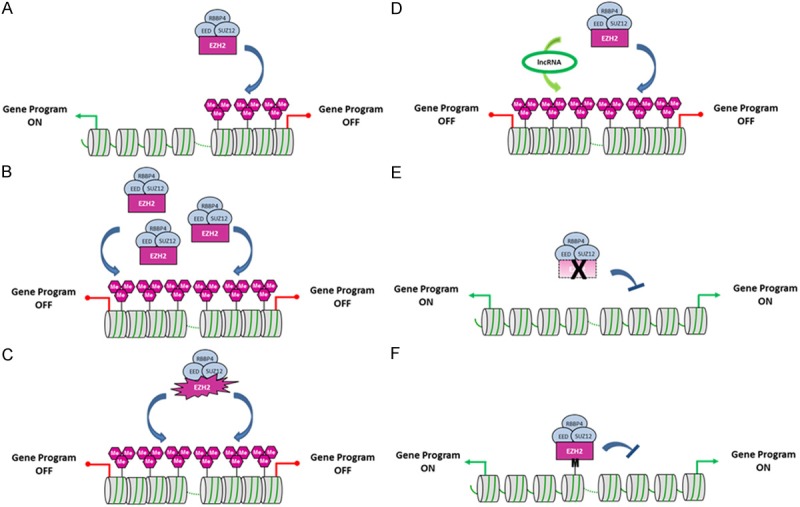
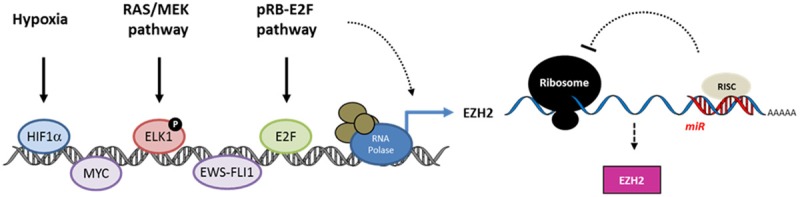
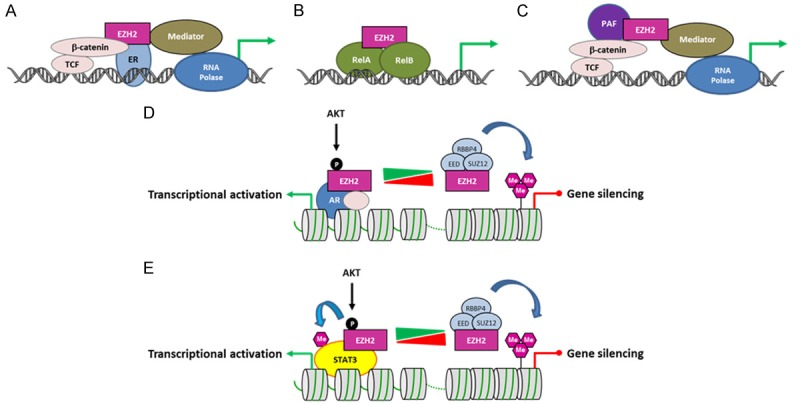
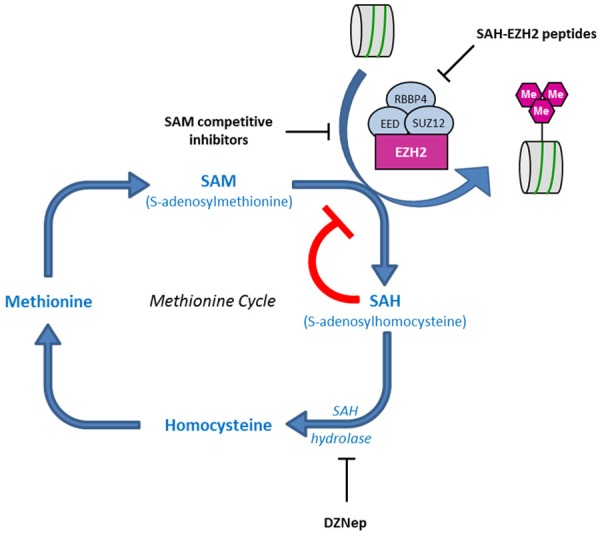
EZH2 is the catalytic subunit of Polycomb Repressor Complex 2 (PRC2) which catalyzes methylation of histone H3 at lysine 27 (H3K27me) and mediates gene silencing of target genes via local chromatin reorganization. Numerous evidences show that EZH2 plays a critical role in cancer initiation, progression and metastasis, as well as in cancer stem cell biology. Indeed, EZH2 dysregulation alters gene expression programs in various cancer types. The molecular mechanisms responsible for EZH2 alteration appear to be diverse and depending on the type of cancer. Furthermore, accumulating evidences indicate that EZH2 could also act as a PRC2-independent transcriptional activator in cancer. In this review, we address the current understanding of the oncogenic role of EZH2, including the mechanisms of EZH2 dysregulation in cancer and progresses in therapeutic approaches targeting EZH2.
Keywords: EZH2; cancer; cancer stem cells; chromatin modification; histone lysine methylation; polycomb repression.
Figures
References
-
- Kennison JA. The Polycomb and trithorax group proteins of Drosophila: trans-regulators of homeotic gene function. Annu Rev Genet. 1995;29:289–303. - PubMed
-
- Schuettenbruger B, Chourrout D, Vervoort M, Leblanc B, Cavalli G. Genome regulation by Polycomb and Trithorax proteins. Cell. 2007;128:735–745. - PubMed
-
- Jürgens G. A group of genes controlling the special expression of the bithorax complex in Drosophila. Nature. 1985;316:153–155.
-
- Yu BD, Hess JL, Horning SE, Brown GAJ, Korsmeyer SJ. Altered Hox expression and segmental identity in Mll-mutant mice. Nature. 1995;378:505–508. - PubMed
-
- Akasaka T, Kanno M, Balling R, Mieza MA, Taniguchi M, Koseki H. A role for mel-18, a Polycomb group-related vertebrate gene, during the anteroposterior specification of the axial skeleton. Development. 1996;122:1513–1522. - PubMed
Publication types
LinkOut - more resources
Full Text Sources