

Coupled local translation and degradation regulate growth cone collapse
- PMID: 25901863
- PMCID: PMC4408908
- DOI: 10.1038/ncomms7888
Coupled local translation and degradation regulate growth cone collapse
Abstract
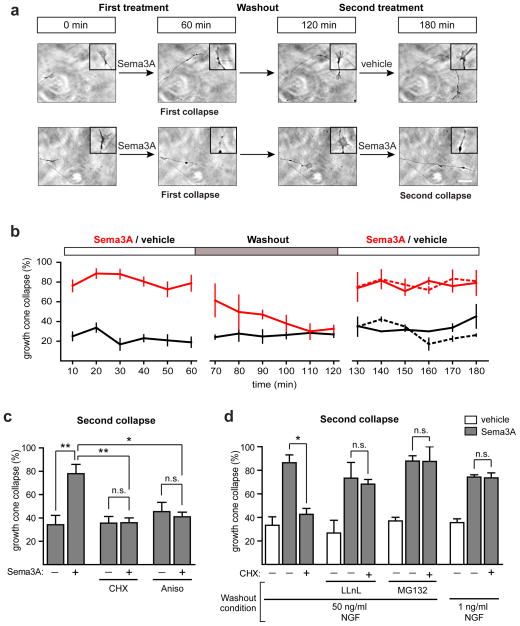
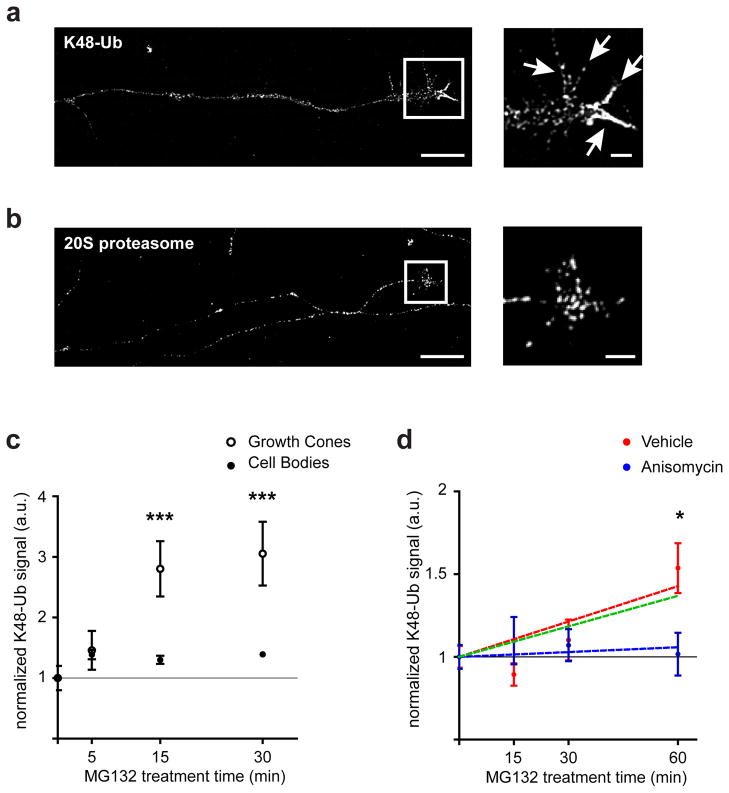
Local translation mediates axonal responses to Semaphorin3A (Sema3A) and other guidance cues. However, only a subset of the axonal proteome is locally synthesized, whereas most proteins are trafficked from the soma. The reason why only specific proteins are locally synthesized is unknown. Here we show that local protein synthesis and degradation are linked events in growth cones. We find that growth cones exhibit high levels of ubiquitination and that local signalling pathways trigger the ubiquitination and degradation of RhoA, a mediator of Sema3A-induced growth cone collapse. Inhibition of RhoA degradation is sufficient to remove the protein-synthesis requirement for Sema3A-induced growth cone collapse. In addition to RhoA, we find that locally translated proteins are the main targets of the ubiquitin-proteasome system in growth cones. Thus, local protein degradation is a major feature of growth cones and creates a requirement for local translation to replenish proteins needed to maintain growth cone responses.
Conflict of interest statement
The authors declare no competing financial interests.
Figures




Similar articles
-
Local translation of RhoA regulates growth cone collapse.Nature. 2005 Aug 18;436(7053):1020-1024. doi: 10.1038/nature03885. Nature. 2005. PMID: 16107849 Free PMC article.
-
Protein synthesis in distal axons is not required for growth cone responses to guidance cues.J Neurosci. 2009 Jan 21;29(3):638-52. doi: 10.1523/JNEUROSCI.3845-08.2009. J Neurosci. 2009. PMID: 19158291 Free PMC article.
-
Semaphorin-3A Promotes Degradation of Fragile X Mental Retardation Protein in Growth Cones via the Ubiquitin-Proteasome Pathway.Front Neural Circuits. 2020 Feb 28;14:5. doi: 10.3389/fncir.2020.00005. eCollection 2020. Front Neural Circuits. 2020. PMID: 32184710 Free PMC article.
-
Growth cones integrate signaling from multiple guidance cues.J Histochem Cytochem. 2003 Apr;51(4):435-44. doi: 10.1177/002215540305100405. J Histochem Cytochem. 2003. PMID: 12642622 Review.
-
Regulation of growth cone actin filaments by guidance cues.J Neurobiol. 2004 Jan;58(1):92-102. doi: 10.1002/neu.10282. J Neurobiol. 2004. PMID: 14598373 Review.
Cited by
-
Phototriggered protein syntheses by using (7-diethylaminocoumarin-4-yl)methoxycarbonyl-caged aminoacyl tRNAs.Nat Commun. 2016 Aug 17;7:12501. doi: 10.1038/ncomms12501. Nat Commun. 2016. PMID: 27530762 Free PMC article.
-
Recent progress in translational engineered in vitro models of the central nervous system.Brain. 2020 Dec 5;143(11):3181-3213. doi: 10.1093/brain/awaa268. Brain. 2020. PMID: 33020798 Free PMC article. Review.
-
Division of labor in the growth cone by DSCR1.J Cell Biol. 2016 May 23;213(4):407-9. doi: 10.1083/jcb.201605012. J Cell Biol. 2016. PMID: 27216257 Free PMC article.
-
Local translation in neuronal compartments: how local is local?EMBO Rep. 2017 May;18(5):693-711. doi: 10.15252/embr.201744045. Epub 2017 Apr 12. EMBO Rep. 2017. PMID: 28404606 Free PMC article. Review.
-
Multiple Copies of microRNA Binding Sites in Long 3'UTR Variants Regulate Axonal Translation.Cells. 2023 Jan 6;12(2):233. doi: 10.3390/cells12020233. Cells. 2023. PMID: 36672174 Free PMC article.
References
-
- Campbell DS, Holt CE. Chemotropic responses of retinal growth cones mediated by rapid local protein synthesis and degradation. Neuron. 2001;32:1013–1026. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
