

Streptozotocin Stimulates the Ion Channel TRPA1 Directly: INVOLVEMENT OF PEROXYNITRITE
- PMID: 25903127
- PMCID: PMC4463460
- DOI: 10.1074/jbc.M115.644476
Streptozotocin Stimulates the Ion Channel TRPA1 Directly: INVOLVEMENT OF PEROXYNITRITE
Abstract
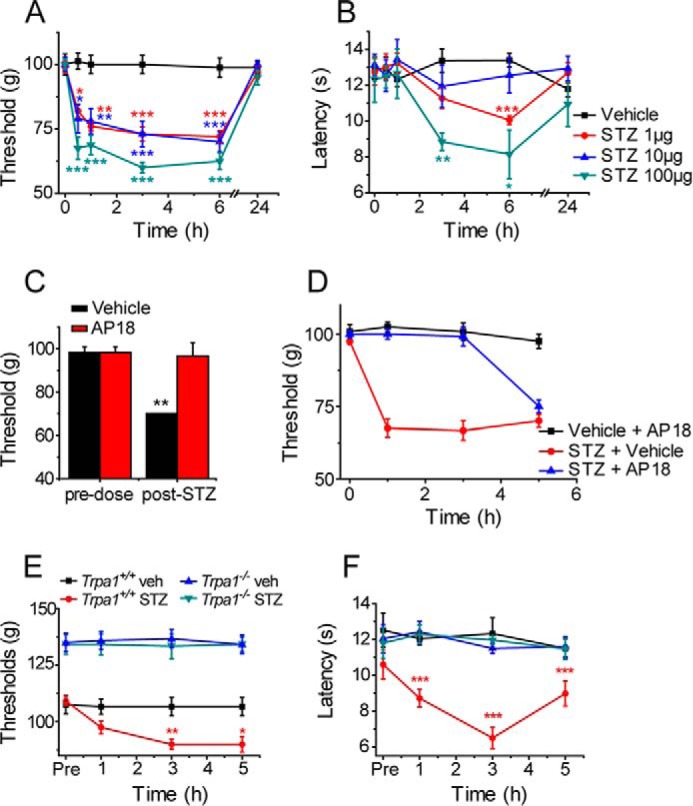
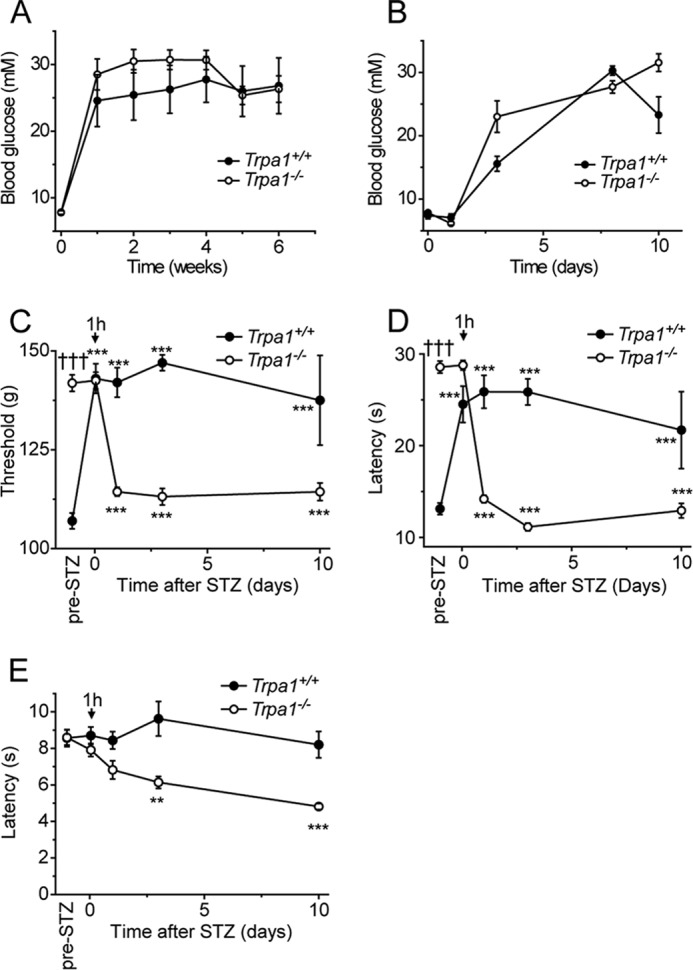
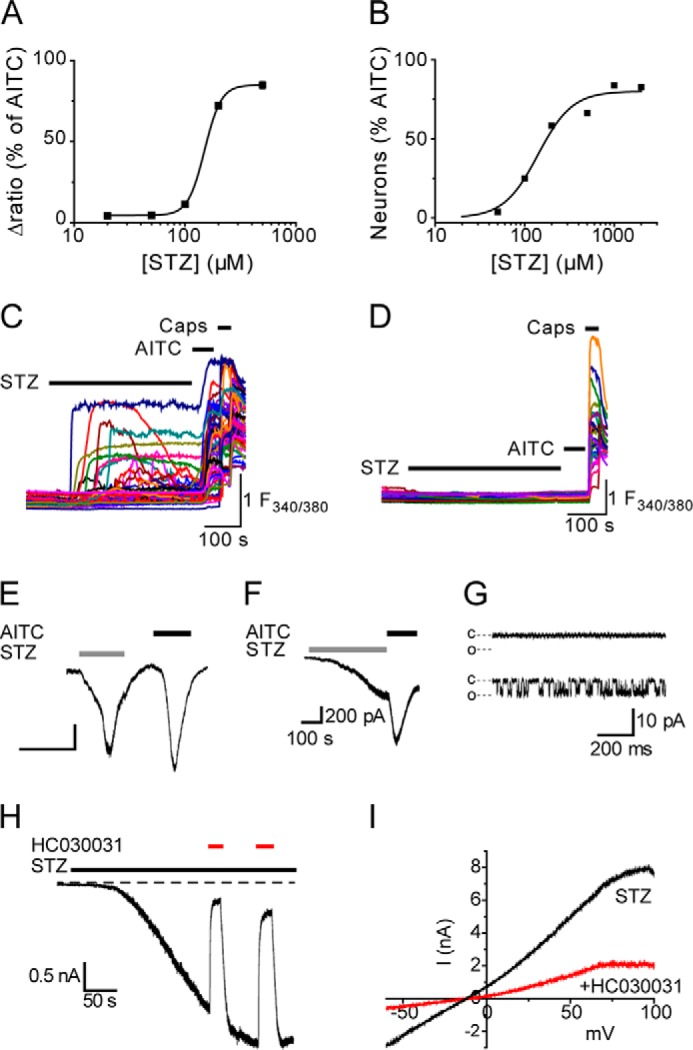
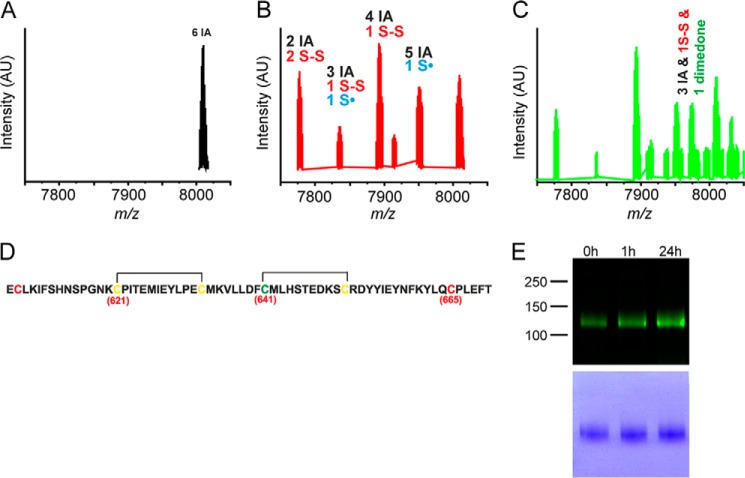
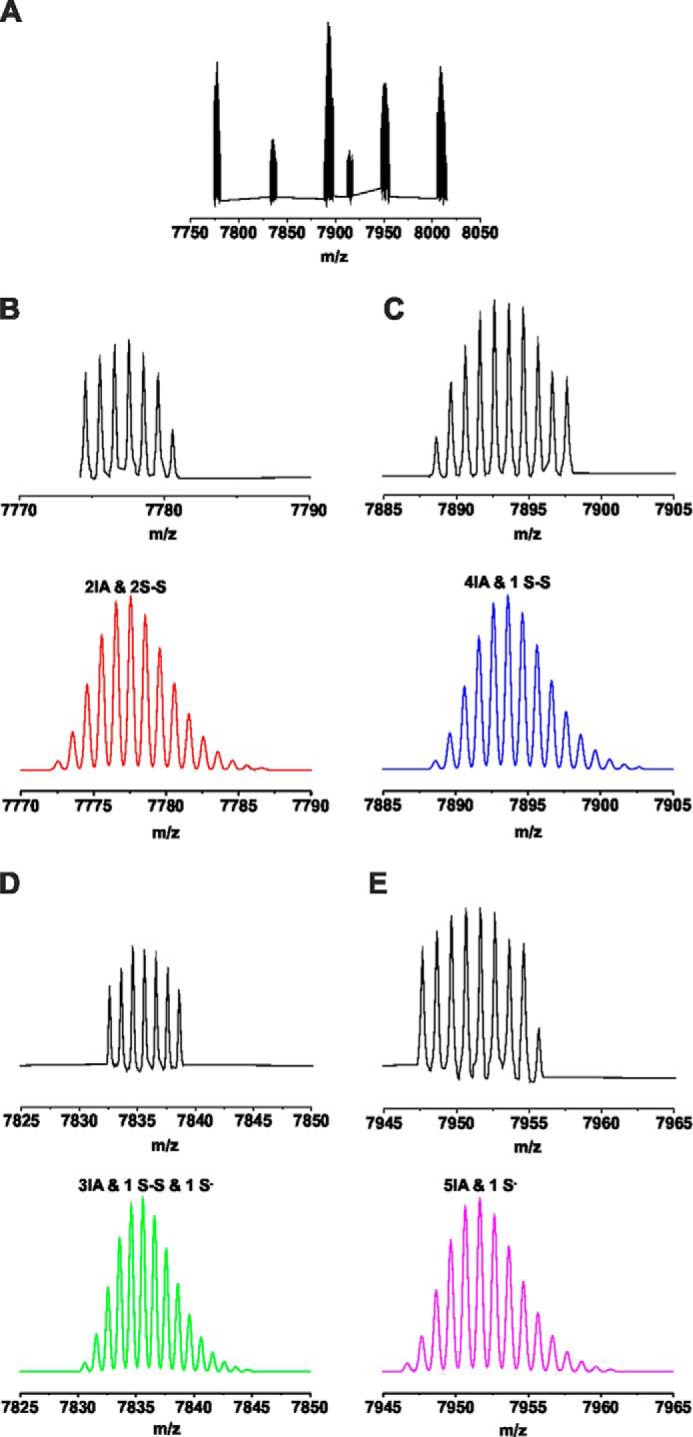
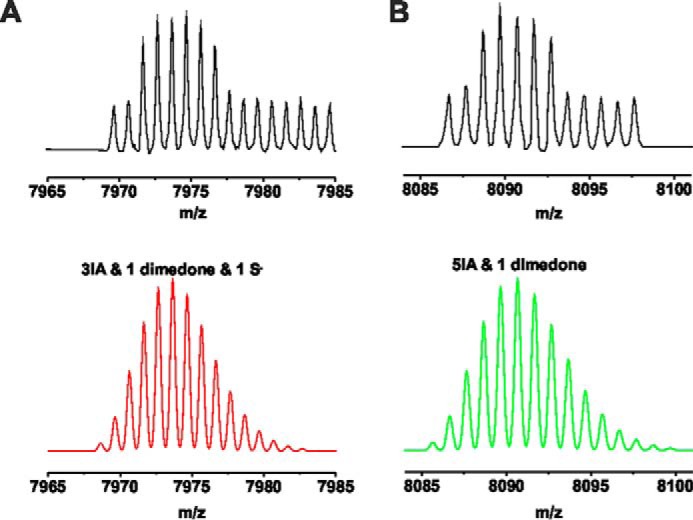
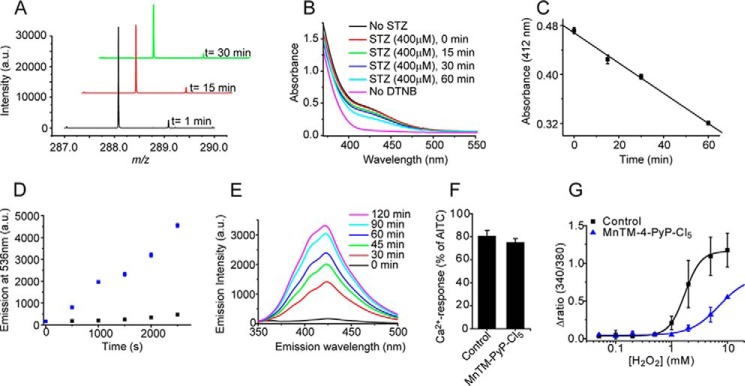
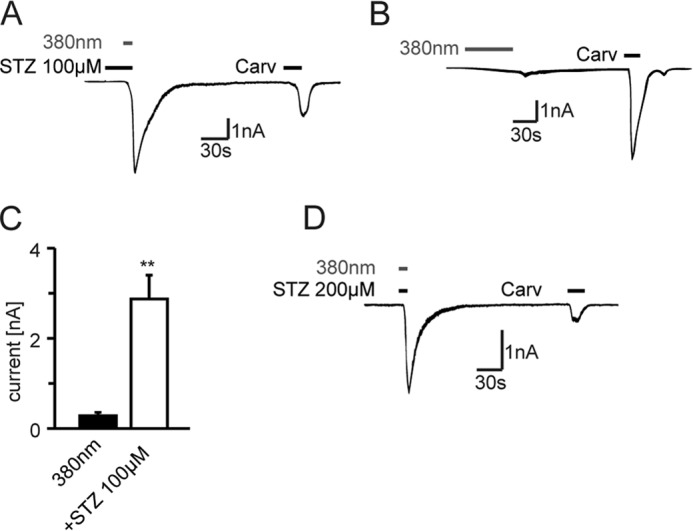
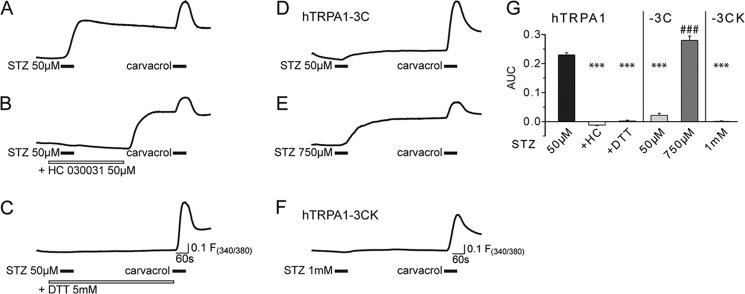
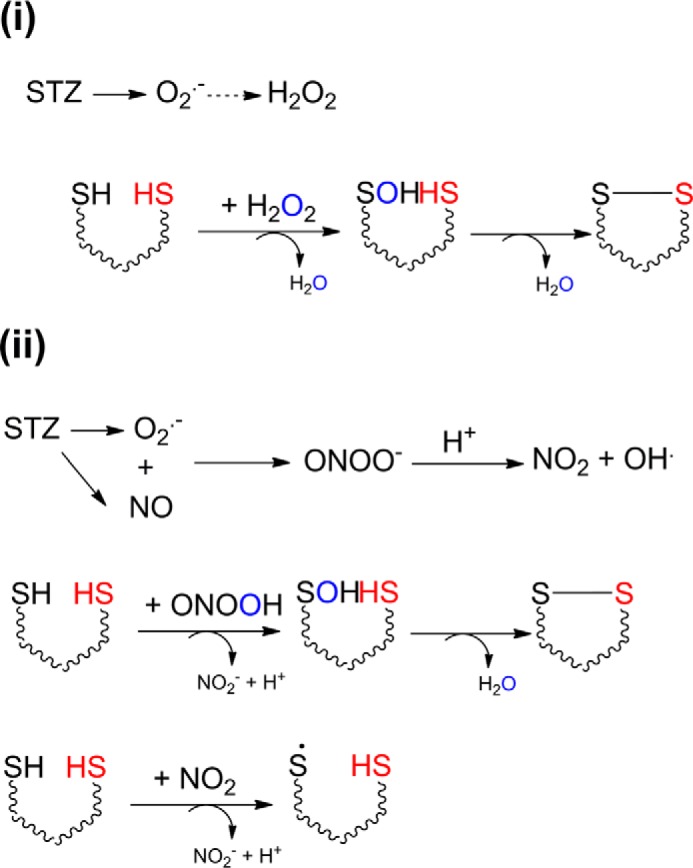
Streptozotocin (STZ)-induced diabetes is the most commonly used animal model of diabetes. Here, we have demonstrated that intraplantar injections of low dose STZ evoked acute polymodal hypersensitivities in mice. These hypersensitivities were inhibited by a TRPA1 antagonist and were absent in TRPA1-null mice. In wild type mice, systemic STZ treatment (180 mg/kg) evoked a loss of cold and mechanical sensitivity within an hour of injection, which lasted for at least 10 days. In contrast, Trpa1(-/-) mice developed mechanical, cold, and heat hypersensitivity 24 h after STZ. The TRPA1-dependent sensory loss produced by STZ occurs before the onset of diabetes and may thus not be readily distinguished from the similar sensory abnormalities produced by the ensuing diabetic neuropathy. In vitro, STZ activated TRPA1 in isolated sensory neurons, TRPA1 cell lines, and membrane patches. Mass spectrometry studies revealed that STZ oxidizes TRPA1 cysteines to disulfides and sulfenic acids. Furthermore, incubation of tyrosine with STZ resulted in formation of dityrosine, suggesting formation of peroxynitrite. Functional analysis of TRPA1 mutants showed that cysteine residues that were oxidized by STZ were important for TRPA1 responsiveness to STZ. Our results have identified oxidation of TRPA1 cysteine residues, most likely by peroxynitrite, as a novel mechanism of action of STZ. Direct stimulation of TRPA1 complicates the interpretation of results from STZ models of diabetic sensory neuropathy and strongly argues that more refined models of diabetic neuropathy should replace the use of STZ.
Keywords: diabetes; dorsal root ganglia; pain; reactive nitrogen species; reactive oxygen species; streptozotocin; transient receptor potential channels (TRP channels).
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Forbes J. M., Cooper M. E. (2013) Mechanisms of diabetic complications. Physiol. Rev. 93, 137–188 - PubMed
-
- Vincent A. M., Callaghan B. C., Smith A. L., Feldman E. L. (2011) Diabetic neuropathy: cellular mechanisms as therapeutic targets. Nat. Rev. Neurol. 7, 573–583 - PubMed
-
- Dworkin R. H., O'Connor A. B., Backonja M., Farrar J. T., Finnerup N. B., Jensen T. S., Kalso E. A., Loeser J. D., Miaskowski C., Nurmikko T. J., Portenoy R. K., Rice A. S., Stacey B. R., Treede R. D., Turk D. C., Wallace M. S. (2007) Pharmacologic management of neuropathic pain: evidence-based recommendations. Pain 132, 237–251 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
