

Involvement of histaminergic inputs in the jaw-closing reflex arc
- PMID: 25904711
- PMCID: PMC4468966
- DOI: 10.1152/jn.00515.2014
Involvement of histaminergic inputs in the jaw-closing reflex arc
Abstract
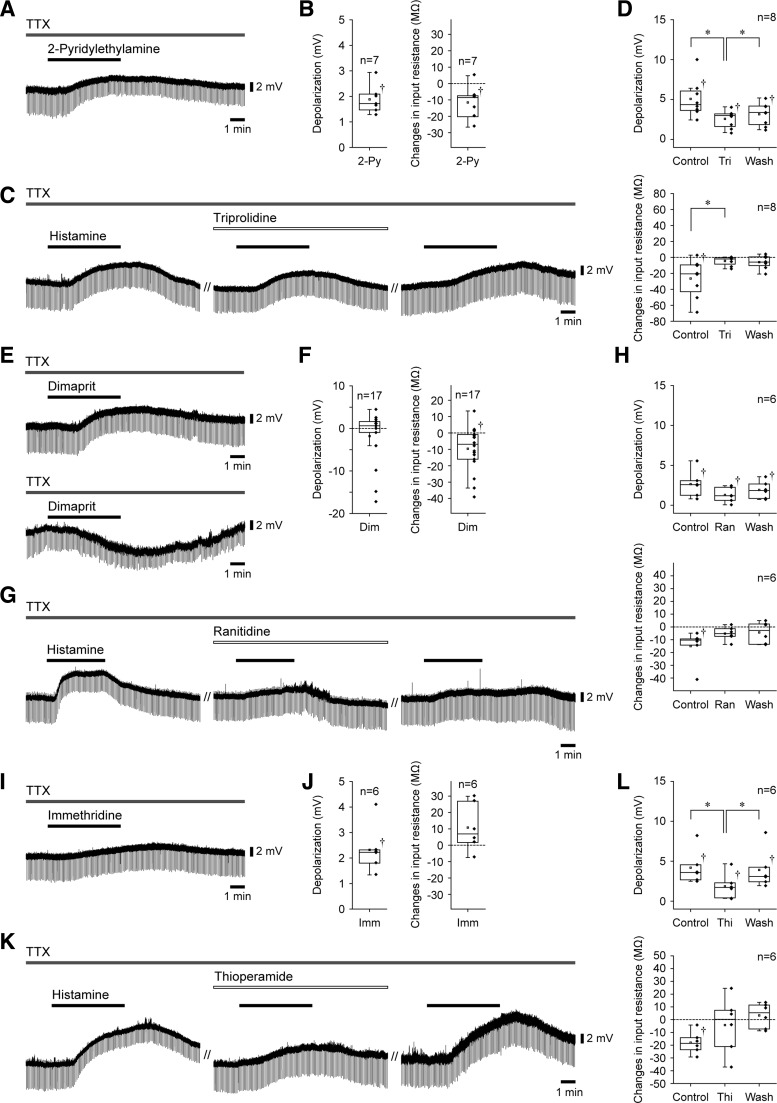
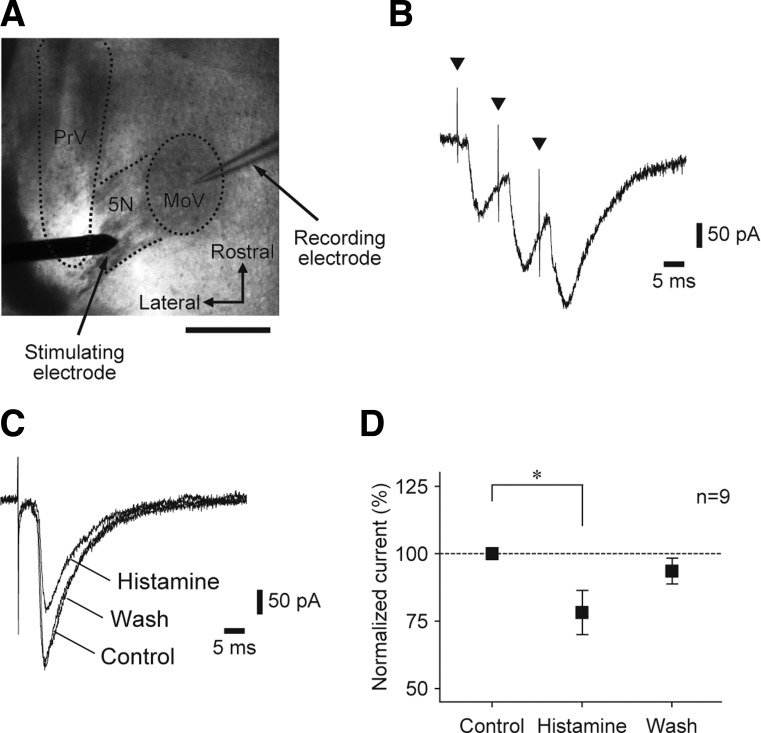
Histamine receptors are densely expressed in the mesencephalic trigeminal nucleus (MesV) and trigeminal motor nucleus. However, little is known about the functional roles of neuronal histamine in controlling oral-motor activity. Thus, using the whole-cell recording technique in brainstem slice preparations from Wistar rats aged between postnatal days 7 and 13, we investigated the effects of histamine on the MesV neurons innervating the masseter muscle spindles and masseter motoneurons (MMNs) that form a reflex arc for the jaw-closing reflex. Bath application of histamine (100 μM) induced membrane depolarization in both MesV neurons and MMNs in the presence of tetrodotoxin, whereas histamine decreased and increased the input resistance in MesV neurons and MMNs, respectively. The effects of histamine on MesV neurons and MMNs were mimicked by an H1 receptor agonist, 2-pyridylethylamine (100 μM). The effects of an H2 receptor agonist, dimaprit (100 μM), on MesV neurons were inconsistent, whereas MMNs were depolarized without changes in the input resistance. An H3 receptor agonist, immethridine (100 μM), also depolarized both MesV neurons and MMNs without changing the input resistance. Histamine reduced the peak amplitude of postsynaptic currents (PSCs) in MMNs evoked by stimulation of the trigeminal motor nerve (5N), which was mimicked by 2-pyridylethylamine but not by dimaprit or immethridine. Moreover, 2-pyridylethylamine increased the failure rate of PSCs evoked by minimal stimulation and the paired-pulse ratio. These results suggest that histaminergic inputs to MesV neurons through H1 receptors are involved in the suppression of the jaw-closing reflex although histamine depolarizes MesV neurons and/or MMNs.
Keywords: histamine; jaw-closing reflex; masseter motoneuron; mesencephalic trigeminal nucleus; patch clamp.
Copyright © 2015 the American Physiological Society.
Figures






Similar articles
-
Serotonin1B receptor-mediated presynaptic inhibition of proprioceptive sensory inputs to jaw-closing motoneurons.Brain Res Bull. 2019 Jul;149:260-267. doi: 10.1016/j.brainresbull.2019.05.001. Epub 2019 May 7. Brain Res Bull. 2019. PMID: 31075304
-
Excitatory effect of histamine on rat spinal motoneurons by activation of both H₁ and H₂ receptors in vitro.J Neurosci Res. 2012 Jan;90(1):132-42. doi: 10.1002/jnr.22730. Epub 2011 Sep 15. J Neurosci Res. 2012. PMID: 21922515
-
Properties of synaptic transmission from the reticular formation dorsal to the facial nucleus to trigeminal motoneurons during early postnatal development in rats.Neuroscience. 2010 Mar 31;166(3):1008-22. doi: 10.1016/j.neuroscience.2009.12.065. Epub 2010 Jan 6. Neuroscience. 2010. PMID: 20060035
-
Projection and synaptic connectivity of trigeminal mesencephalic nucleus neurons controlling jaw reflexes.J Oral Sci. 2017;59(2):177-182. doi: 10.2334/josnusd.16-0845. J Oral Sci. 2017. PMID: 28637975 Review.
-
Histaminergic transmission in the mammalian brain.Physiol Rev. 1991 Jan;71(1):1-51. doi: 10.1152/physrev.1991.71.1.1. Physiol Rev. 1991. PMID: 1846044 Review. No abstract available.
Cited by
-
Histamine 1 receptor-Gβγ-cAMP/PKA-CFTR pathway mediates the histamine-induced resetting of the suprachiasmatic circadian clock.Mol Brain. 2016 May 6;9(1):49. doi: 10.1186/s13041-016-0227-1. Mol Brain. 2016. PMID: 27153809 Free PMC article.
References
-
- Appenteng K, Girdlestone D. Transneuronal transport of wheat germ agglutinin-conjugated horseradish peroxidase into trigeminal interneurones of the rat. J Comp Neurol 258: 387–396, 1987. - PubMed
-
- Arrang JM, Garbarg M, Schwartz JC. Autoregulation of histamine release in brain by presynaptic H3-receptors. Neuroscience 15: 553–562, 1985. - PubMed
-
- Arrang JM, Garbarg M, Schwartz JC. Autoinhibition of histamine synthesis mediated by presynaptic H3-receptors. Neuroscience 23: 149–157, 1987. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources