

Spinal dopaminergic projections control the transition to pathological pain plasticity via a D1/D5-mediated mechanism
- PMID: 25904784
- PMCID: PMC4405552
- DOI: 10.1523/JNEUROSCI.3481-14.2015
Spinal dopaminergic projections control the transition to pathological pain plasticity via a D1/D5-mediated mechanism
Abstract
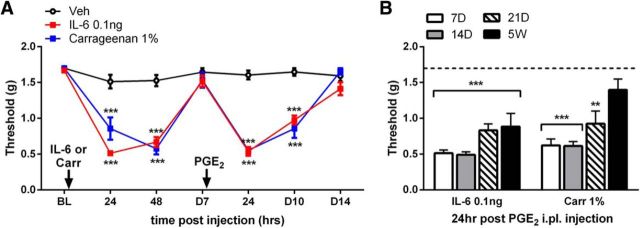
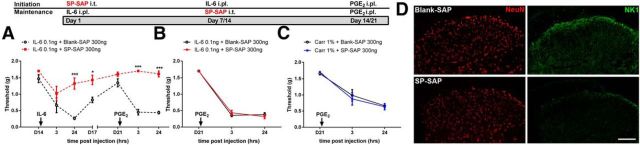
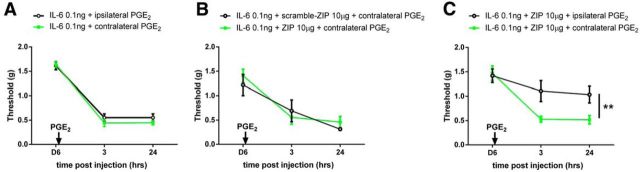
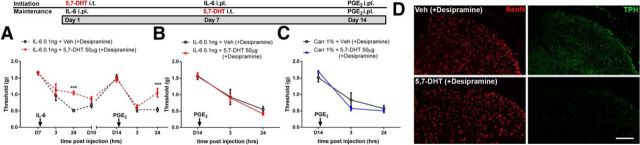
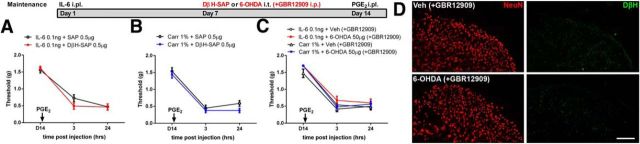
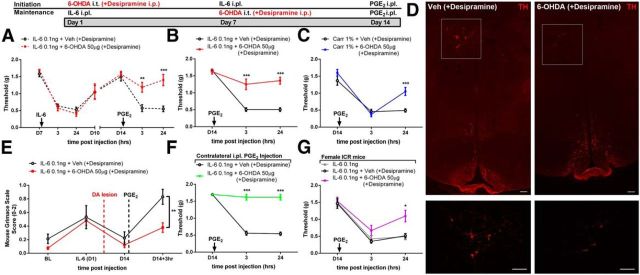
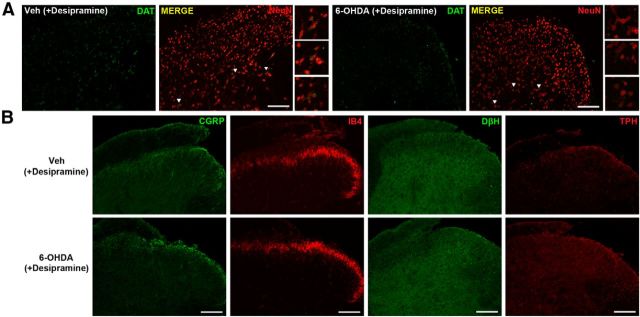
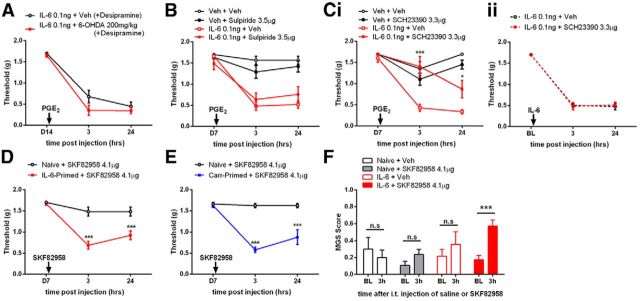
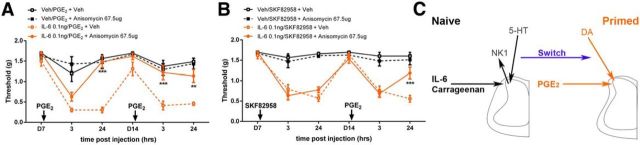
The mechanisms that lead to the maintenance of chronic pain states are poorly understood, but their elucidation could lead to new insights into how pain becomes chronic and how it can potentially be reversed. We investigated the role of spinal dorsal horn neurons and descending circuitry in plasticity mediating a transition to pathological pain plasticity suggesting the presence of a chronic pain state using hyperalgesic priming. We found that when dorsal horn neurokinin 1 receptor-positive neurons or descending serotonergic neurons were ablated before hyperalgesic priming, IL-6- and carrageenan-induced mechanical hypersensitivity was impaired, and subsequent prostaglandin E2 (PGE2) response was blunted. However, when these neurons were lesioned after the induction of priming, they had no effect on the PGE2 response, reflecting differential mechanisms driving plasticity in a primed state. In stark contrast, animals with a spinally applied dopaminergic lesion showed intact IL-6- and carrageenan-induced mechanical hypersensitivity, but the subsequent PGE2 injection failed to cause mechanical hypersensitivity. Moreover, ablating spinally projecting dopaminergic neurons after the resolution of the IL-6- or carrageenan-induced response also reversed the maintenance of priming as assessed through mechanical hypersensitivity and the mouse grimace scale. Pharmacological antagonism of spinal dopamine D1/D5 receptors reversed priming, whereas D1/D5 agonists induced mechanical hypersensitivity exclusively in primed mice. Strikingly, engagement of D1/D5 coupled with anisomycin in primed animals reversed a chronic pain state, consistent with reconsolidation-like effects in the spinal dorsal horn. These findings demonstrate a novel role for descending dopaminergic neurons in the maintenance of pathological pain plasticity.
Keywords: chronic pain; descending modulation; dopamine; nociceptive plasticity; reconsolidation; substance P.
Copyright © 2015 the authors 0270-6474/15/356307-11$15.00/0.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources