

Diet impact on mitochondrial bioenergetics and dynamics
- PMID: 25904870
- PMCID: PMC4389347
- DOI: 10.3389/fphys.2015.00109
Diet impact on mitochondrial bioenergetics and dynamics
Abstract
Diet induced obesity is associated with impaired mitochondrial function and dynamic behavior. Mitochondria are highly dynamic organelles and the balance in fusion/fission is strictly associated with their bioenergetics. Fusion processes are associated with the optimization of mitochondrial function, whereas fission processes are associated with the removal of damaged mitochondria. In diet-induced obesity, impaired mitochondrial function and increased fission processes were found in liver and skeletal muscle. Diverse dietary fat sources differently affect mitochondrial dynamics and bioenergetics. In contrast to saturated fatty acids, omega 3 polyunsaturated fatty acids induce fusion processes and improve mitochondrial function. Moreover, the pro-longevity effect of caloric restriction has been correlated with changes in mitochondrial dynamics leading to decreased cell oxidative injury. Noteworthy, emerging findings revealed an important role for mitochondrial dynamics within neuronal populations involved in central regulation of body energy balance. In conclusion, mitochondrial dynamic processes with their strict interconnection with mitochondrial bioenergetics are involved in energy balance and diet impact on metabolic tissues.
Keywords: caloric restriction; dietary fat; energy balance; mitochondrial fission; mitochondrial fusion.
Figures
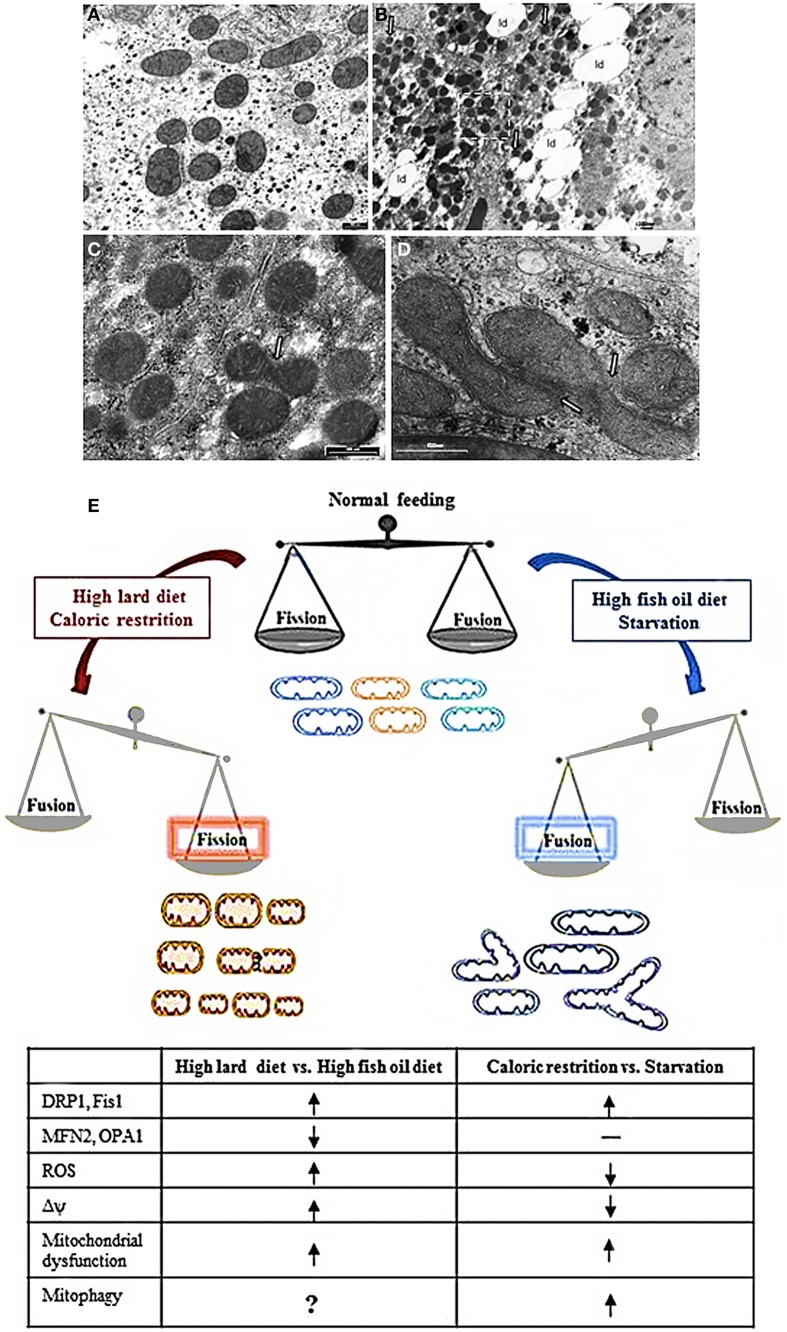
 = increase,
= increase,  = decrease; —= no changes.
= decrease; —= no changes.References
-
- Bach D., Naon D., Pich S., Soriano F. X., Vega N., Rieusset J., et al. . (2005). Expression of Mfn2, the Charcot-Marie-Tooth neuropathy type 2A gene, in human skeletal muscle: effects of type 2 diabetes, obesity, weight loss, and the regulatory role of tumor necrosis factor alpha and interleukin-6. Diabetes 54, 2685–2693. 10.2337/diabetes.54.9.2685 - DOI - PubMed
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources