

Time and Demand are Two Critical Dimensions of Immunometabolism: The Process of Macrophage Activation and the Pentose Phosphate Pathway
- PMID: 25904920
- PMCID: PMC4389563
- DOI: 10.3389/fimmu.2015.00164
Time and Demand are Two Critical Dimensions of Immunometabolism: The Process of Macrophage Activation and the Pentose Phosphate Pathway
Abstract
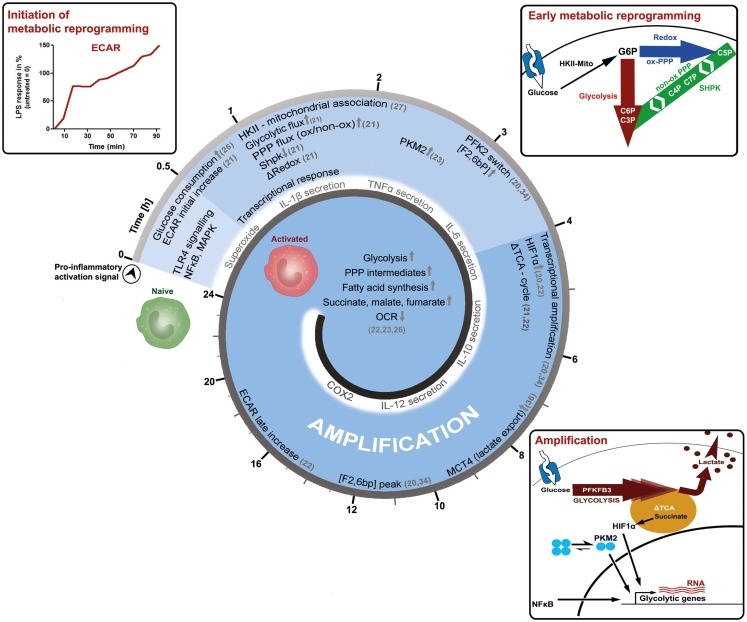
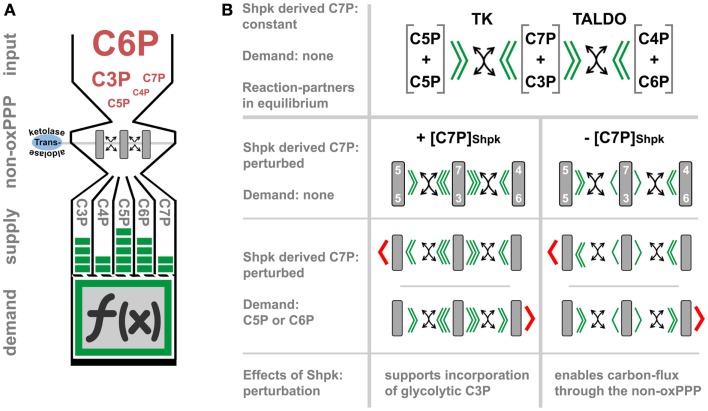
A process is a function of time; in immunometabolism, this is reflected by the stepwise adaptation of metabolism to sustain the bio-energetic demand of an immune-response in its various states and shades. This perspective article starts by presenting an early attempt to investigate the physiology of inflammation, in order to illustrate one of the basic concepts of immunometabolism, wherein an adapted metabolism of infiltrating immune cells affects tissue function and inflammation. We then focus on the process of macrophage activation and aim to delineate the factor time within the current molecular context of metabolic-rewiring important for adapting primary carbohydrate metabolism. In the last section, we will provide information on how the pentose phosphate pathway may be of importance to provide both nucleotide precursors and redox-equivalents, and speculate how carbon-scrambling events in the non-oxidative pentose phosphate pathway might be regulated within cells by demand. We conclude that the adapted metabolism of inflammation is specific in respect to the effector-function and appears as a well-orchestrated event, dynamic by nature, and based on a functional interplay of signaling- and metabolic-pathways.
Keywords: immunometabolism; inflammation; macrophage activation; metabolic reprograming; pentose phosphate pathway; primary carbohydrate metabolism; sedoheptulose kinase; time and demand.
Figures
Similar articles
-
Sedoheptulose kinase regulates cellular carbohydrate metabolism by sedoheptulose 7-phosphate supply.Biochem Soc Trans. 2013 Apr;41(2):674-80. doi: 10.1042/BST20120354. Biochem Soc Trans. 2013. PMID: 23514175 Review.
-
The sedoheptulose kinase CARKL controls T-cell cytokine outputs and migration by promoting metabolic reprogramming.Discov Immunol. 2024 Nov 19;3(1):kyae016. doi: 10.1093/discim/kyae016. eCollection 2024. Discov Immunol. 2024. PMID: 39669692 Free PMC article.
-
Adipose tissue at the nexus of systemic and cellular immunometabolism.Semin Immunol. 2016 Oct;28(5):431-440. doi: 10.1016/j.smim.2016.09.005. Epub 2016 Oct 24. Semin Immunol. 2016. PMID: 27789087 Review.
-
The significance of sedoheptulose 1,7-bisphosphate in the metabolism and regulation of the pentose pathway in liver.Biochem Int. 1985 Oct;11(4):599-610. Biochem Int. 1985. PMID: 4084320
-
A model for the role of the proline-linked pentose-phosphate pathway in phenolic phytochemical bio-synthesis and mechanism of action for human health and environmental applications.Asia Pac J Clin Nutr. 2004;13(1):1-24. Asia Pac J Clin Nutr. 2004. PMID: 15003910 Review.
Cited by
-
Reversal of β-Amyloid-Induced Microglial Toxicity In Vitro by Activation of Fpr2/3.Oxid Med Cell Longev. 2020 Jun 13;2020:2139192. doi: 10.1155/2020/2139192. eCollection 2020. Oxid Med Cell Longev. 2020. PMID: 32617132 Free PMC article.
-
Hexokinase 1 cellular localization regulates the metabolic fate of glucose.Mol Cell. 2022 Apr 7;82(7):1261-1277.e9. doi: 10.1016/j.molcel.2022.02.028. Epub 2022 Mar 18. Mol Cell. 2022. PMID: 35305311 Free PMC article.
-
The emerging roles of metabolism in the crosstalk between breast cancer cells and tumor-associated macrophages.Int J Biol Sci. 2023 Sep 18;19(15):4915-4930. doi: 10.7150/ijbs.86039. eCollection 2023. Int J Biol Sci. 2023. PMID: 37781517 Free PMC article. Review.
-
Identification of Discriminating Metabolic Pathways and Metabolites in Human PBMCs Stimulated by Various Pathogenic Agents.Front Physiol. 2018 Feb 27;9:139. doi: 10.3389/fphys.2018.00139. eCollection 2018. Front Physiol. 2018. PMID: 29535640 Free PMC article.
-
Metabolic Crossroad Between Macrophages and Cancer Cells: Overview of Hepatocellular Carcinoma.Biomedicines. 2024 Nov 25;12(12):2684. doi: 10.3390/biomedicines12122684. Biomedicines. 2024. PMID: 39767591 Free PMC article. Review.
References
-
- Levene PA, Meyer GM. The action of leucocytes on glucose. J Biol Chem (1912) 12:361–70.
-
- Warburg O. Stoffwechsel der Tumoren. Berlin: Springer; (1926).
-
- Bakker A. Einige uebereinstimmungen im stoffwechsel der carcinomzellen und exsudatleukocyten. Klin Wochenschr (1927) 6:252–410.1007/BF01710710 - DOI
-
- Fleischmann W, Kubowitz F. Ueber den stoffwechsel der leukocyten. Biochem Ztschr (1927) 181:395.
-
- Kempner W, Peschel E. Stoffwechsel der entzuendung. Ztschr f klin Med (1930) 114:439–55.
Publication types
LinkOut - more resources
Full Text Sources
Other Literature Sources