

Essential role for the ATG4B protease and autophagy in bleomycin-induced pulmonary fibrosis
- PMID: 25906080
- PMCID: PMC4502665
- DOI: 10.1080/15548627.2015.1034409
Essential role for the ATG4B protease and autophagy in bleomycin-induced pulmonary fibrosis
Abstract
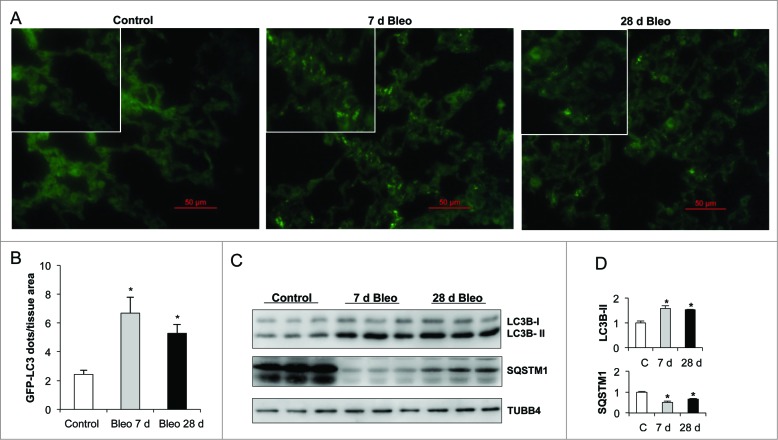
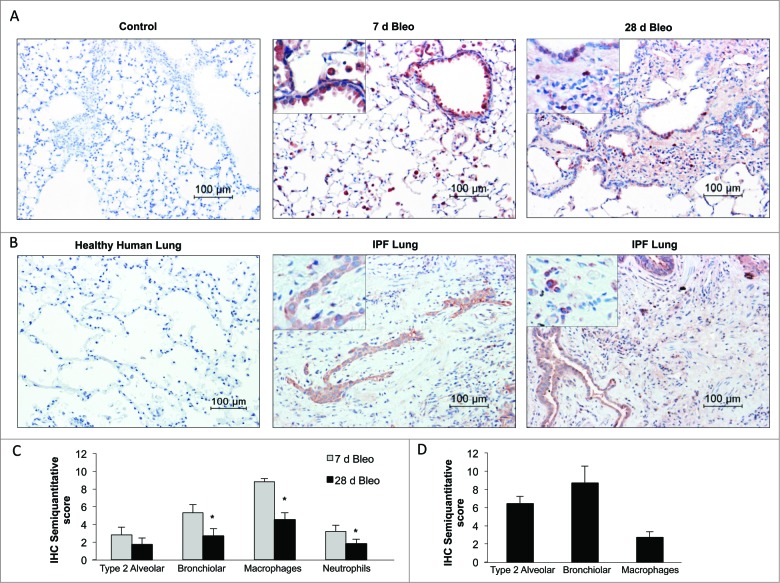
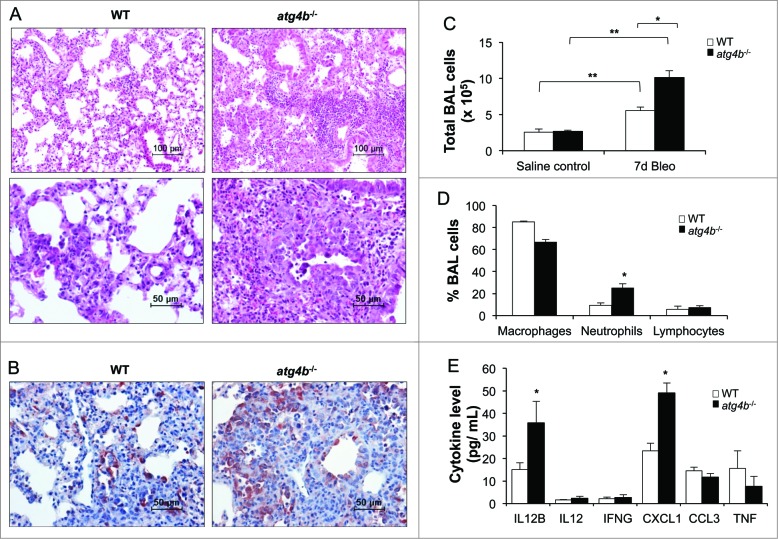
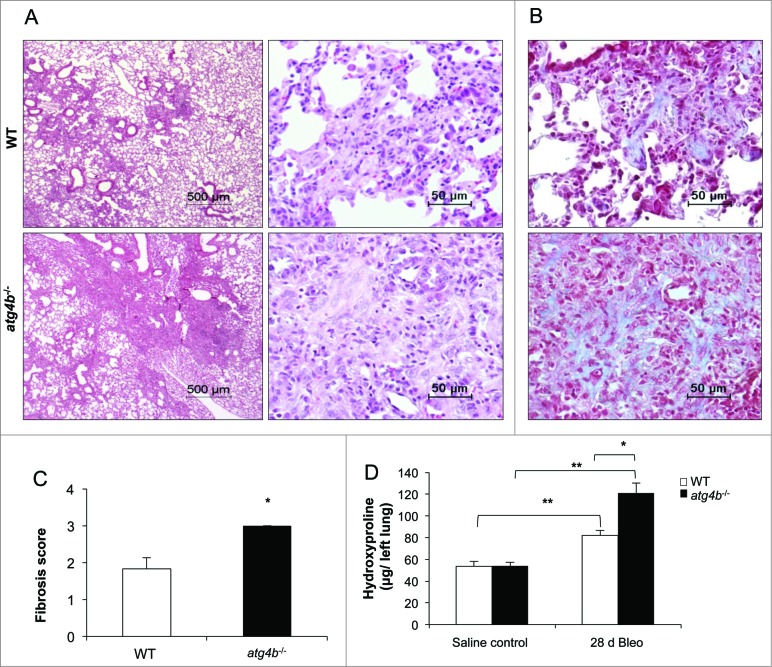
Autophagy is a critical cellular homeostatic process that controls the turnover of damaged organelles and proteins. Impaired autophagic activity is involved in a number of diseases, including idiopathic pulmonary fibrosis suggesting that altered autophagy may contribute to fibrogenesis. However, the specific role of autophagy in lung fibrosis is still undefined. In this study, we show for the first time, how autophagy disruption contributes to bleomycin-induced lung fibrosis in vivo using an Atg4b-deficient mouse as a model. Atg4b-deficient mice displayed a significantly higher inflammatory response at 7 d after bleomycin treatment associated with increased neutrophilic infiltration and significant alterations in proinflammatory cytokines. Likewise, we found that Atg4b disruption resulted in augmented apoptosis affecting predominantly alveolar and bronchiolar epithelial cells. At 28 d post-bleomycin instillation Atg4b-deficient mice exhibited more extensive and severe fibrosis with increased collagen accumulation and deregulated extracellular matrix-related gene expression. Together, our findings indicate that the ATG4B protease and autophagy play a crucial role protecting epithelial cells against bleomycin-induced stress and apoptosis, and in the regulation of the inflammatory and fibrotic responses.
Keywords: ACTA2, actin, α 2, smooth muscle, aorta; ATG3, autophagy-related 3; ATG4B; ATG4B, autophagy-related 4B; ATG5, autophagy-related 5; ATG7, autophagy-related 7; ATG9B, autophagy-related 9B; BAX, BCL2-associated X protein; CASP3, caspase 3, apoptosis-related cysteine peptidase; CAV1, caveolin 1, caveolae protein, 22kDa; CCL3, chemokine (C-C motif) ligand 3; CXCL1, chemokine (C-X-C motif) ligand 1 (melanoma growth stimulating activity α); CXCR2, chemokine (C-X-C motif) receptor 2; DRAM2, DNA-damage regulated autophagy modulator 2; GFP-LC3B, green fluorescent protein-LC3B; IFNG, interferon, gamma; IL12B, interleukin 12B; IL13, interleukin 13; IPF, idiopathic pulmonary fibrosis; MAP1LC3B/LC3B, microtubule-associated protein 1 light chain 3 β; RELA, v-rel reticuloendotheliosis viral oncogene homolog A; SQSTM1, sequestosome 1; TGFB1, transforming growth factor, β 1; TGFBR2, transforming growth factor, β receptor II (70/80kDa); TNF, tumor necrosis factor; TUBB4, tubulin, β 4, class IV; WT, wild type; autophagin-1; autophagy; cysteine peptidase; epithelial cell; idiopathic pulmonary fibrosis; lung fibrosis.
Figures





References
-
- Selman M, King TE, Pardo A. Idiopathic pulmonary fibrosis: prevailing and evolving hypotheses about its pathogenesis and implications for therapy. Ann Intern Med 2001; 134:136–51; PMID:11177318; http://dx.doi.org/ 10.7326/0003-4819-134-2-200101160-00015 - DOI - PubMed
-
- King TE, Pardo A, Selman M. Idiopathic pulmonary fibrosis. Lancet 2011; 378:1949–61; PMID:21719092; http://dx.doi.org/ 10.1016/S0140-6736(11)60052-4 - DOI - PubMed
-
- Selman M, Pardo A. Idiopathic pulmonary fibrosis: an epithelial/fibroblastic cross-talk disorder. Respir Res 2002; 3:3; PMID:11806838; http://dx.doi.org/ 10.1186/rr175 - DOI - PMC - PubMed
-
- Selman M, Pardo A. Alveolar epithelial cell disintegrity and subsequent activation: a key process in pulmonary fibrosis. Am J RespirCrit Care Med 2012; 186:119–21; http://dx.doi.org/ 10.1164/rccm.201206-0997ED - DOI - PubMed
-
- Selman M, Rojas M, Mora AL, Pardo A. Aging and interstitial lung diseases: unraveling an old forgotten player in the pathogenesis of lung fibrosis. SeminRespirCrit Care Med 2010; 31:607–17 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous