

Presynaptic BK channel localization is dependent on the hierarchical organization of alpha-catulin and dystrobrevin and fine-tuned by CaV2 calcium channels
- PMID: 25907097
- PMCID: PMC4411755
- DOI: 10.1186/s12868-015-0166-2
Presynaptic BK channel localization is dependent on the hierarchical organization of alpha-catulin and dystrobrevin and fine-tuned by CaV2 calcium channels
Abstract
Background: Large conductance, calcium-activated BK channels regulate many important physiological processes, including smooth muscle excitation, hormone release and synaptic transmission. The biological roles of these channels hinge on their unique ability to respond synergistically to both voltage and cytosolic calcium elevations. Because calcium influx is meticulously regulated both spatially and temporally, the localization of BK channels near calcium channels is critical for their proper function. However, the mechanism underlying BK channel localization near calcium channels is not fully understood.
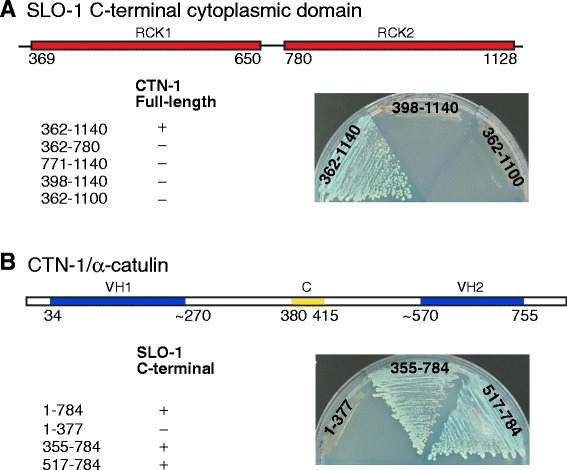
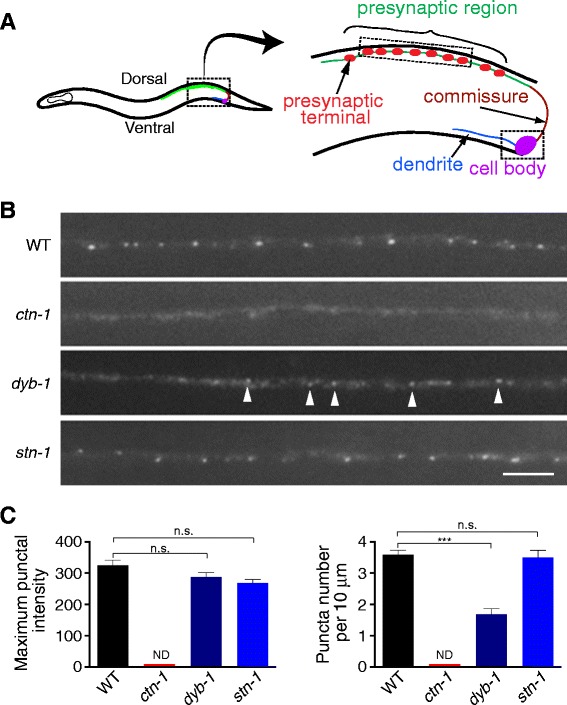
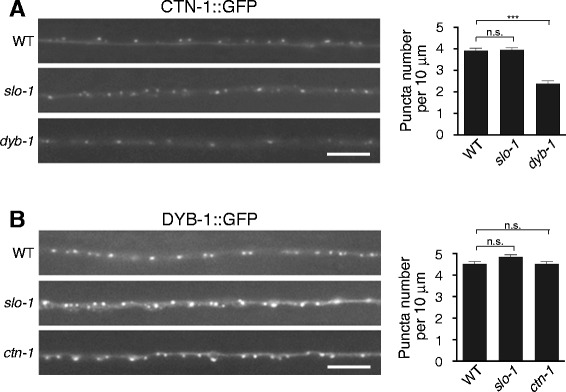
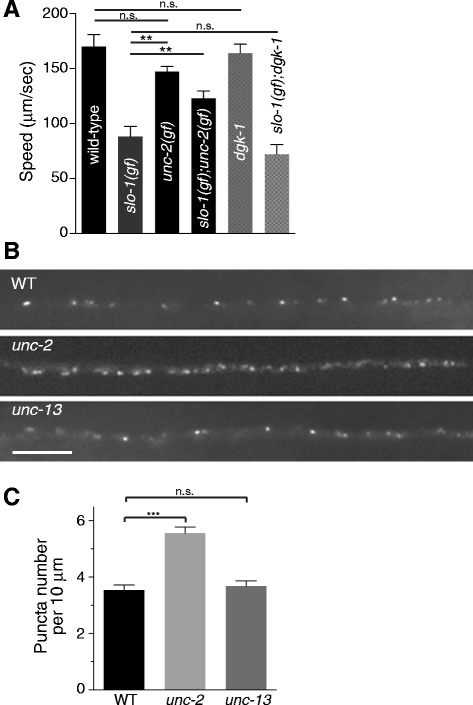
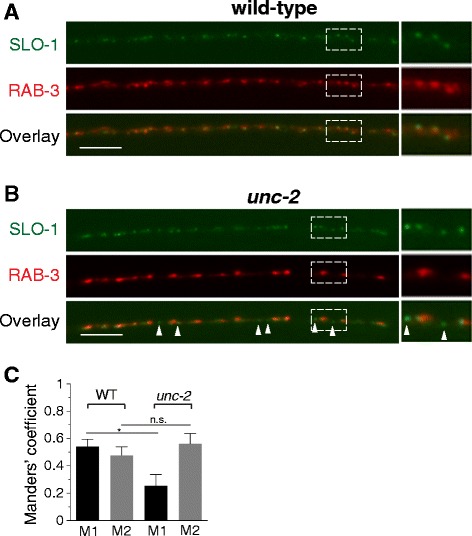
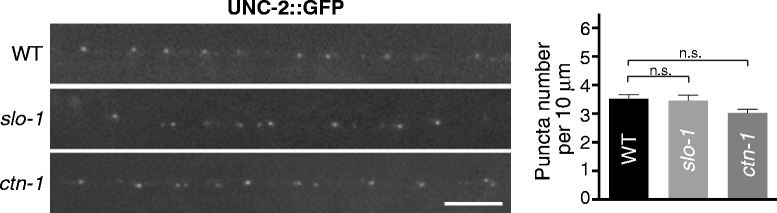
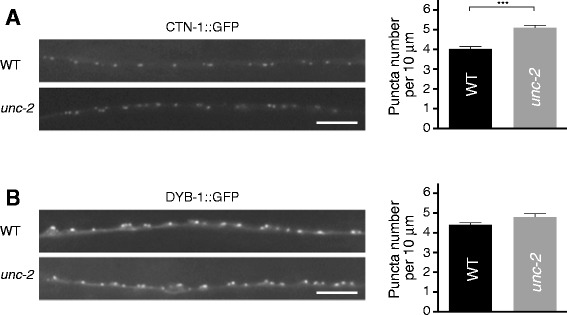
Results: We show here that in C. elegans the localization of SLO-1/BK channels to presynaptic terminals, where UNC-2/CaV2 calcium channels regulate neurotransmitter release, is controlled by the hierarchical organization of CTN-1/α-catulin and DYB-1/dystrobrevin, two proteins that interact with cortical cytoskeletal proteins. CTN-1 organizes a macromolecular SLO-1 channel complex at presynaptic terminals by direct physical interaction. DYB-1 contributes to the maintenance or stabilization of the complex at presynaptic terminals by interacting with CTN-1. We also show that SLO-1 channels are functionally coupled with UNC-2 calcium channels, and that normal localization of SLO-1 to presynaptic terminals requires UNC-2. In the absence of UNC-2, SLO-1 clusters lose the localization specificity, thus accumulating inside and outside of presynaptic terminals. Moreover, CTN-1 is also similarly localized in unc-2 mutants, consistent with the direct interaction between CTN-1 and SLO-1. However, localization of UNC-2 at the presynaptic terminals is not dependent on either CTN-1 or SLO-1. Taken together, our data strongly suggest that the absence of UNC-2 indirectly influences SLO-1 localization via the reorganization of cytoskeletal proteins.
Conclusion: CTN-1 and DYB-1, which interact with cortical cytoskeletal proteins, are required for the presynaptic punctate localization of SLO-1 in a hierarchical manner. In addition, UNC-2 calcium channels indirectly control the fidelity of SLO-1 puncta localization at presynaptic terminals. We suggest that the absence of UNC-2 leads to the reorganization of the cytoskeletal structure that includes CTN-1, which in turn influences SLO-1 puncta localization.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
