

Somatic stem cell heterogeneity: diversity in the blood, skin and intestinal stem cell compartments
- PMID: 25907613
- PMCID: PMC5317203
- DOI: 10.1038/nrm3980
Somatic stem cell heterogeneity: diversity in the blood, skin and intestinal stem cell compartments
Abstract
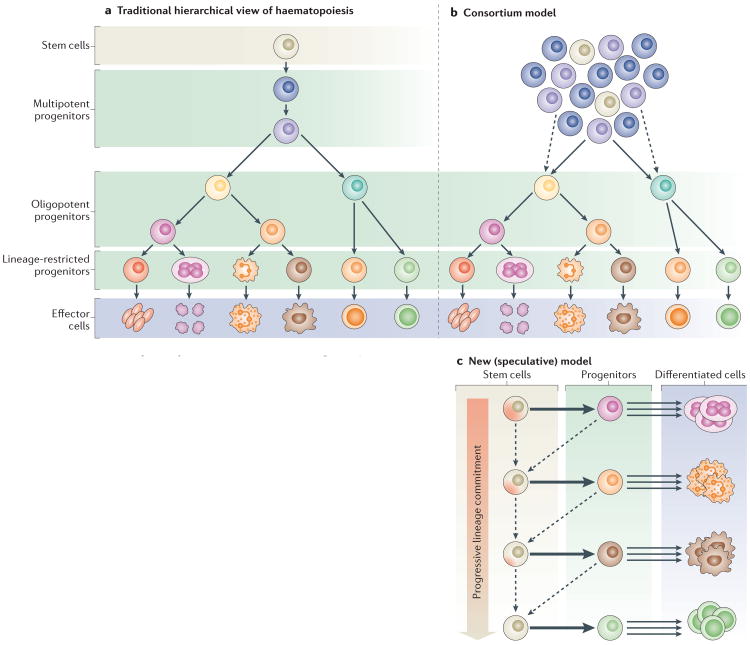
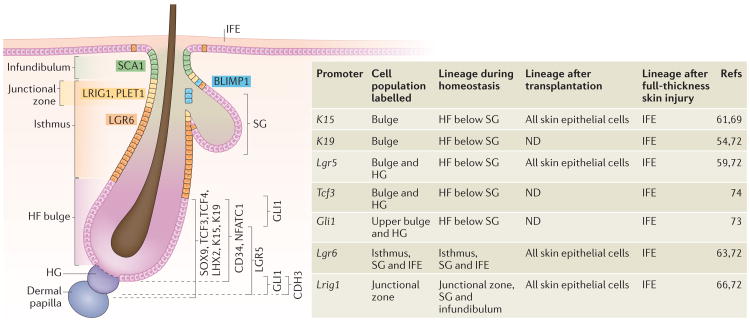
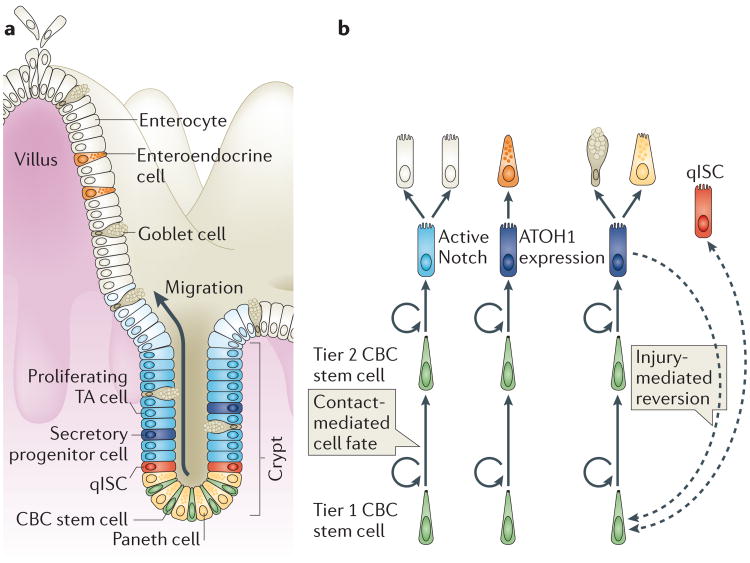
Somatic stem cells replenish many tissues throughout life to repair damage and to maintain tissue homeostasis. Stem cell function is frequently described as following a hierarchical model in which a single master cell undergoes self-renewal and differentiation into multiple cell types and is responsible for most regenerative activity. However, recent data from studies on blood, skin and intestinal epithelium all point to the concomitant action of multiple types of stem cells with distinct everyday roles. Under stress conditions such as acute injury, the surprising developmental flexibility of these stem cells enables them to adapt to diverse roles and to acquire different regeneration capabilities. This paradigm shift raises many new questions about the developmental origins, inter-relationships and molecular regulation of these multiple stem cell types.
Figures
References
-
- Pappenheim A. Virchows Arch Pathol Anat Physiol Klin Med. Vol. 145. in German: 1896. Ueber Entwickelung und Ausbildung der Erythroblasten; pp. 587–643.
-
- Ramalho-Santos M, Willenbring H. On the origin of the term “stem cell”. Cell Stem Cell. 2007;1:35–38. - PubMed
-
- Becker AJ, McCulloch EA, Till JE. Cytological demonstration of the clonal nature of spleen colonies derived from transplanted mouse marrow cells. Nature. 1963;197:452–454. - PubMed
-
- Lemischka IR, Raulet DH, Mulligan RC. Developmental potential and dynamic behavior of hematopoietic stem cells. Cell. 1986;45:917–927. - PubMed
Publication types
MeSH terms
Grants and funding
- R01AR059122/AR/NIAMS NIH HHS/United States
- R01 AR059122/AR/NIAMS NIH HHS/United States
- DK092883/DK/NIDDK NIH HHS/United States
- U01 DK103117/DK/NIDDK NIH HHS/United States
- DK092456/DK/NIDDK NIH HHS/United States
- U24 DK085532/DK/NIDDK NIH HHS/United States
- R01 DK092456/DK/NIDDK NIH HHS/United States
- U01 DK085532/DK/NIDDK NIH HHS/United States
- R01 DK092306/DK/NIDDK NIH HHS/United States
- R01 CA183252/CA/NCI NIH HHS/United States
- DK103117/DK/NIDDK NIH HHS/United States
- R01 DK092883/DK/NIDDK NIH HHS/United States
- CA1428260/CA/NCI NIH HHS/United States
- DK092306/DK/NIDDK NIH HHS/United States
- R56 DK092883/DK/NIDDK NIH HHS/United States
- CA183252/CA/NCI NIH HHS/United States
- R01 CA142826/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
