

The nucleus is an intracellular propagator of tensile forces in NIH 3T3 fibroblasts
- PMID: 25908852
- PMCID: PMC4457156
- DOI: 10.1242/jcs.161703
The nucleus is an intracellular propagator of tensile forces in NIH 3T3 fibroblasts
Abstract
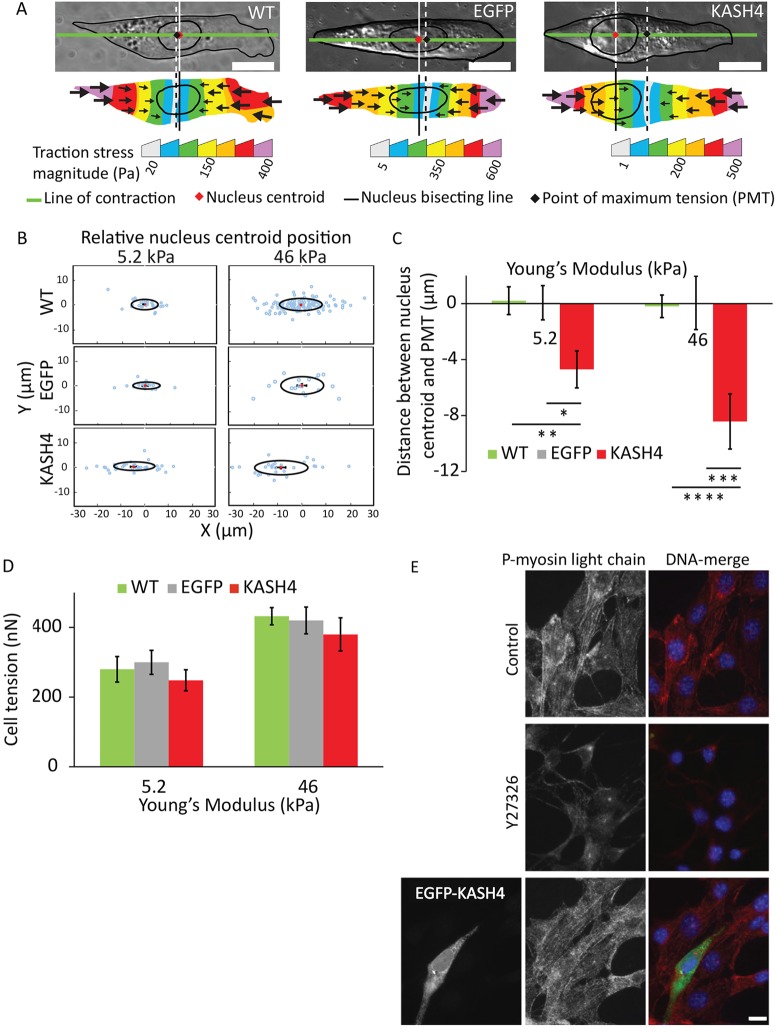
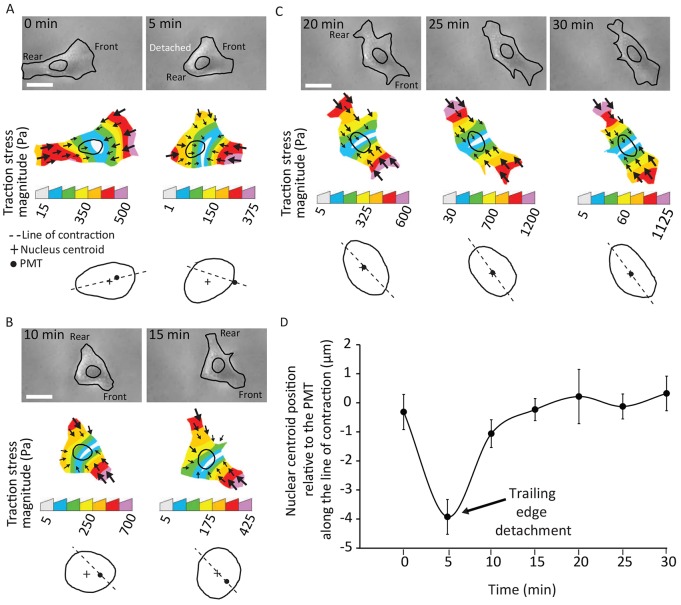
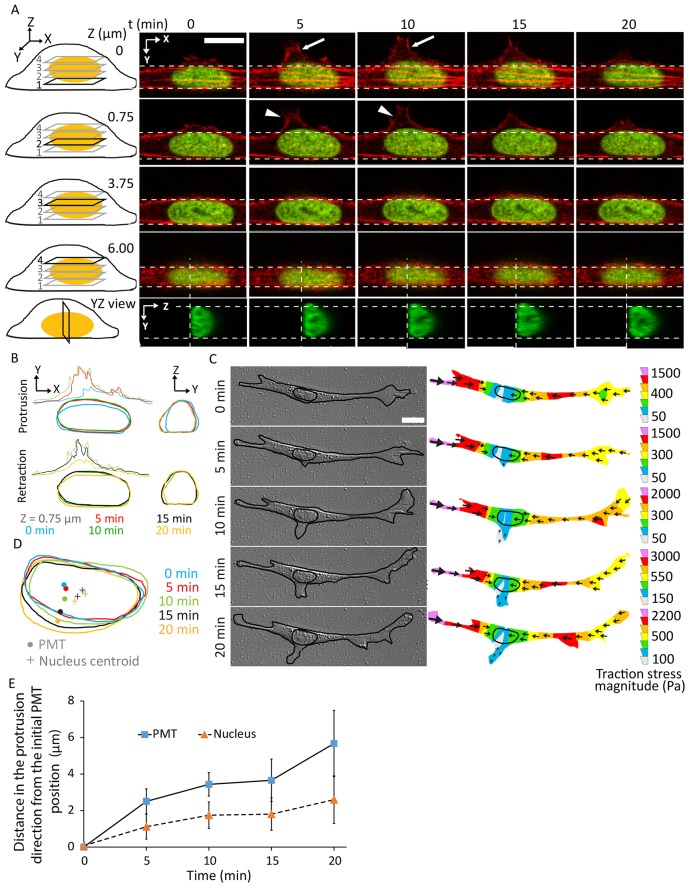
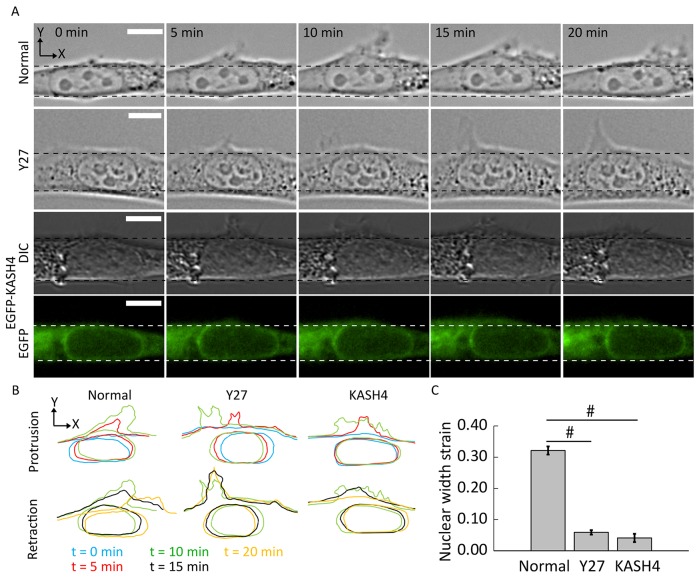
Nuclear positioning is a crucial cell function, but how a migrating cell positions its nucleus is not understood. Using traction-force microscopy, we found that the position of the nucleus in migrating fibroblasts closely coincided with the center point of the traction-force balance, called the point of maximum tension (PMT). Positioning of the nucleus close to the PMT required nucleus-cytoskeleton connections through linker of nucleoskeleton-to-cytoskeleton (LINC) complexes. Although the nucleus briefly lagged behind the PMT following spontaneous detachment of the uropod during migration, the nucleus quickly repositioned to the PMT within a few minutes. Moreover, traction-generating spontaneous protrusions deformed the nearby nucleus surface to pull the nuclear centroid toward the new PMT, and subsequent retraction of these protrusions relaxed the nuclear deformation and restored the nucleus to its original position. We propose that the protruding or retracting cell boundary transmits a force to the surface of the nucleus through the intervening cytoskeletal network connected by the LINC complexes, and that these forces help to position the nucleus centrally and allow the nucleus to efficiently propagate traction forces across the length of the cell during migration.
Keywords: Cytoskeleton; KASH4; LINC complex; Nucleus; Tensile forces; Traction stress.
© 2015. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures
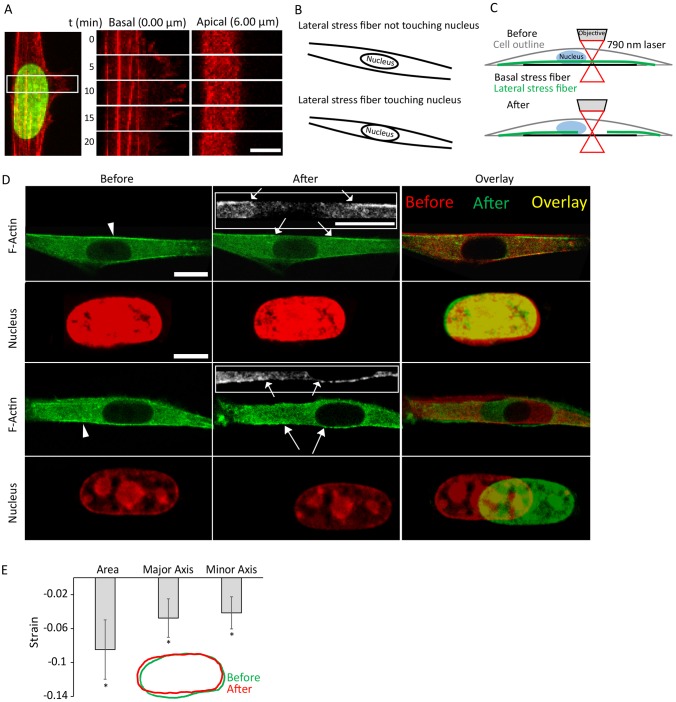
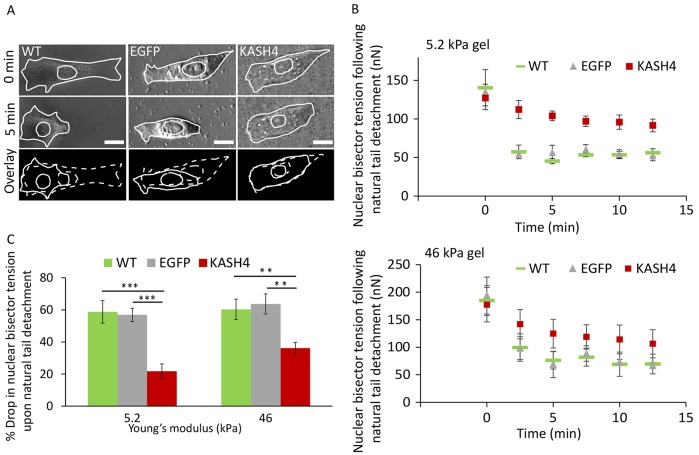
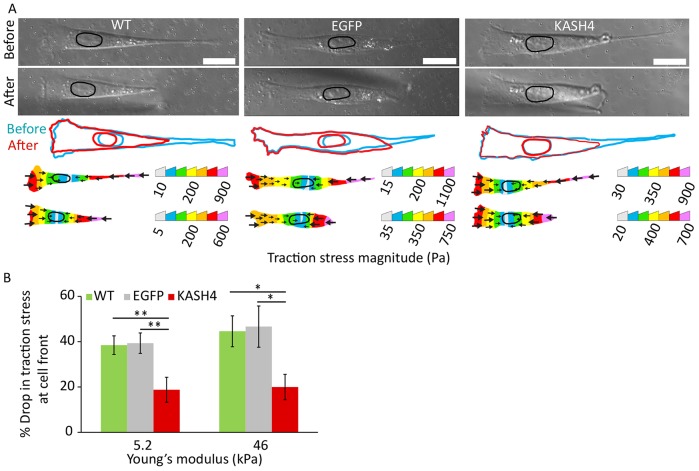
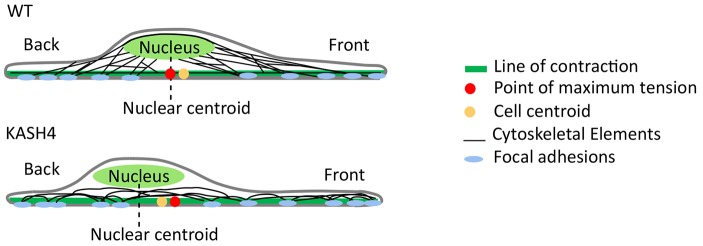
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
