

Induction of stable ER-plasma-membrane junctions by Kv2.1 potassium channels
- PMID: 25908859
- PMCID: PMC4457025
- DOI: 10.1242/jcs.166009
Induction of stable ER-plasma-membrane junctions by Kv2.1 potassium channels
Abstract
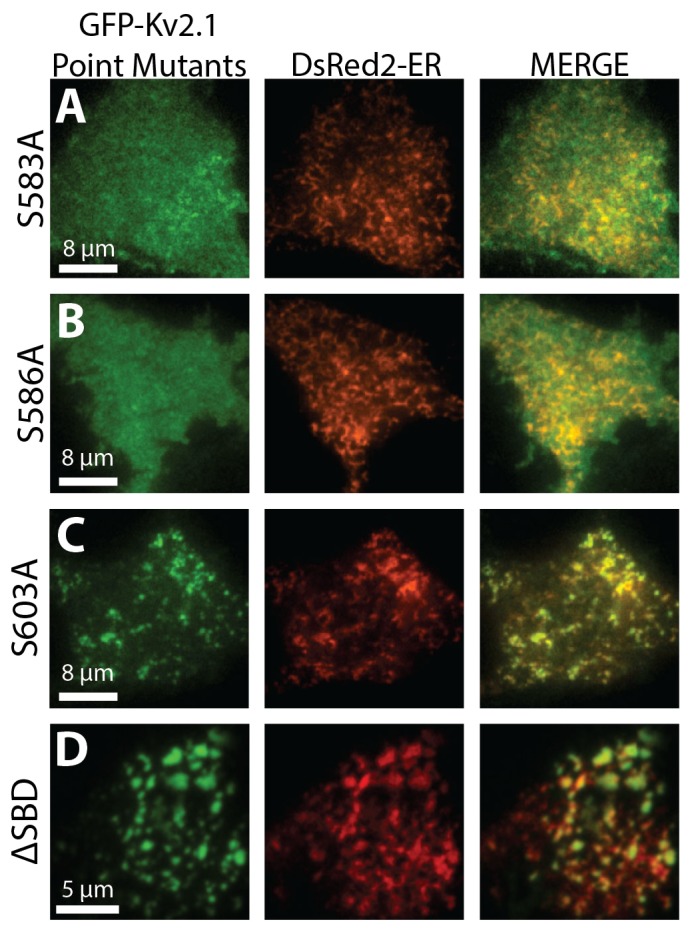
Junctions between cortical endoplasmic reticulum (cER) and the plasma membrane are a subtle but ubiquitous feature in mammalian cells; however, very little is known about the functions and molecular interactions that are associated with neuronal ER-plasma-membrane junctions. Here, we report that Kv2.1 (also known as KCNB1), the primary delayed-rectifier K(+) channel in the mammalian brain, induces the formation of ER-plasma-membrane junctions. Kv2.1 localizes to dense, cell-surface clusters that contain non-conducting channels, indicating that they have a function that is unrelated to membrane-potential regulation. Accordingly, Kv2.1 clusters function as membrane-trafficking hubs, providing platforms for delivery and retrieval of multiple membrane proteins. Using both total internal reflection fluorescence and electron microscopy we demonstrate that the clustered Kv2.1 plays a direct structural role in the induction of stable ER-plasma-membrane junctions in both transfected HEK 293 cells and cultured hippocampal neurons. Glutamate exposure results in a loss of Kv2.1 clusters in neurons and subsequent retraction of the cER from the plasma membrane. We propose Kv2.1-induced ER-plasma-membrane junctions represent a new macromolecular plasma-membrane complex that is sensitive to excitotoxic insult and functions as a scaffolding site for both membrane trafficking and Ca(2+) signaling.
Keywords: Ion channels; Membrane contact sites; Sub-surface cisterns.
© 2015. Published by The Company of Biologists Ltd.
Conflict of interest statement
The authors declare no competing or financial interests.
Figures







References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
