

Toll signals regulate dorsal-ventral patterning and anterior-posterior placement of the embryo in the hemipteran Rhodnius prolixus
- PMID: 25908955
- PMCID: PMC4407881
- DOI: 10.1186/2041-9139-5-38
Toll signals regulate dorsal-ventral patterning and anterior-posterior placement of the embryo in the hemipteran Rhodnius prolixus
Abstract
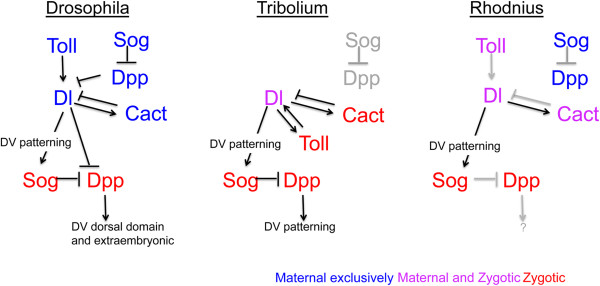
Background: Insect embryonic dorso-ventral patterning depends greatly on two pathways: the Toll pathway and the Bone Morphogenetic Protein pathway. While the relative contribution of each pathway has been investigated in holometabolous insects, their role has not been explored in insects with a hemimetabolous type of development. The hemimetabolous insect Rhodnius prolixus, an important vector of Chagas disease in the Americas, develops from an intermediate germ band and displays complex movements during katatrepsis that are not observed in other orders. However, little is known about the molecular events that regulate its embryogenesis. Here we investigate the expression and function of genes potentially involved in the initial patterning events that establish the embryonic dorso-ventral axis in this hemipteran.
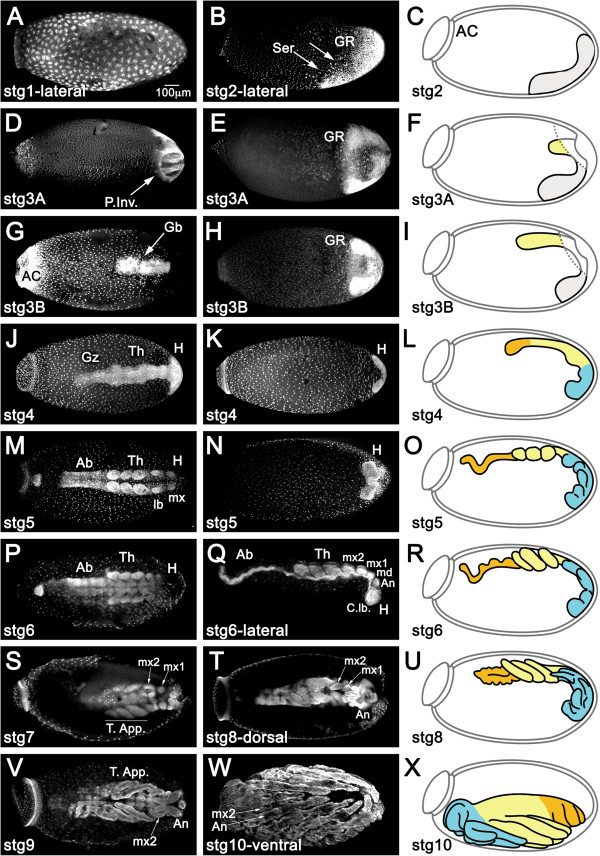
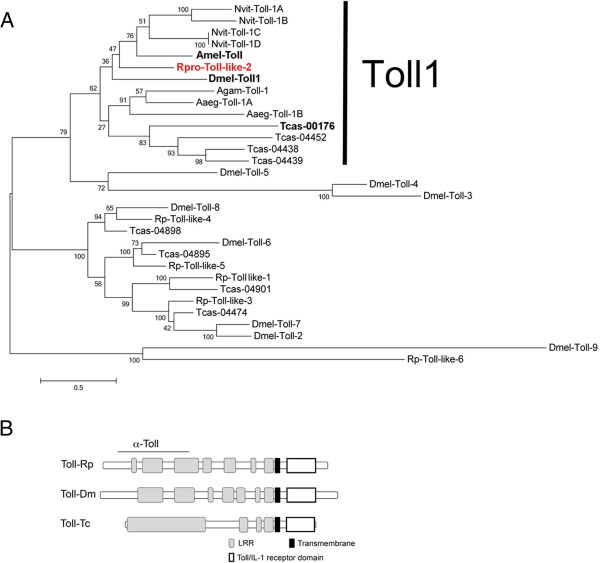
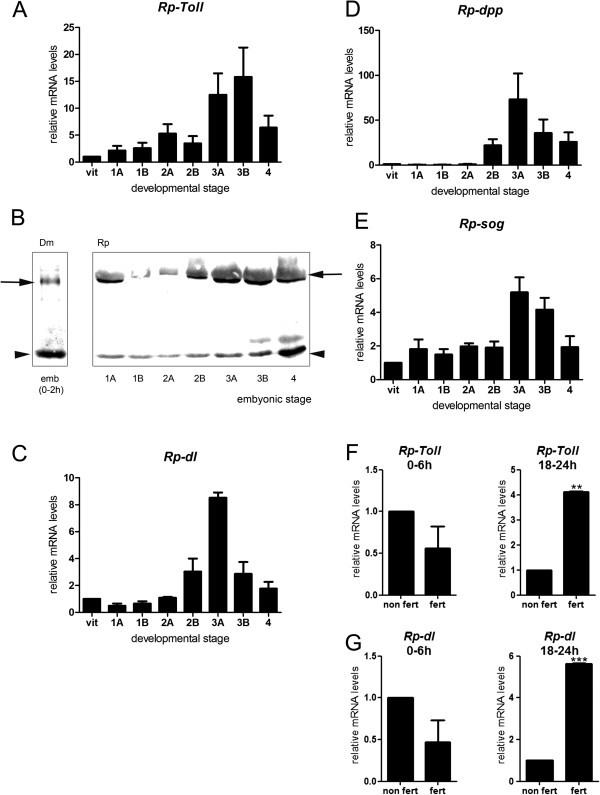
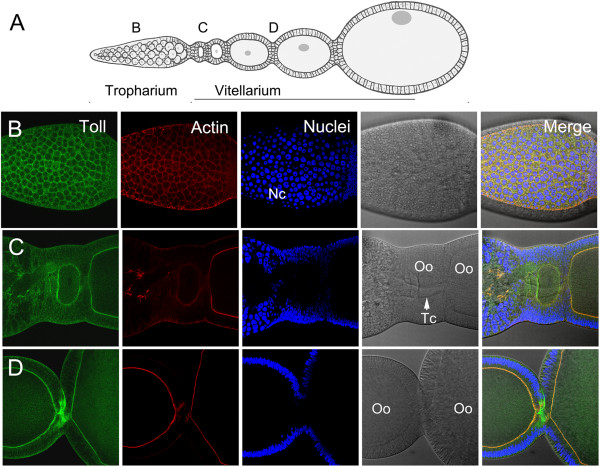
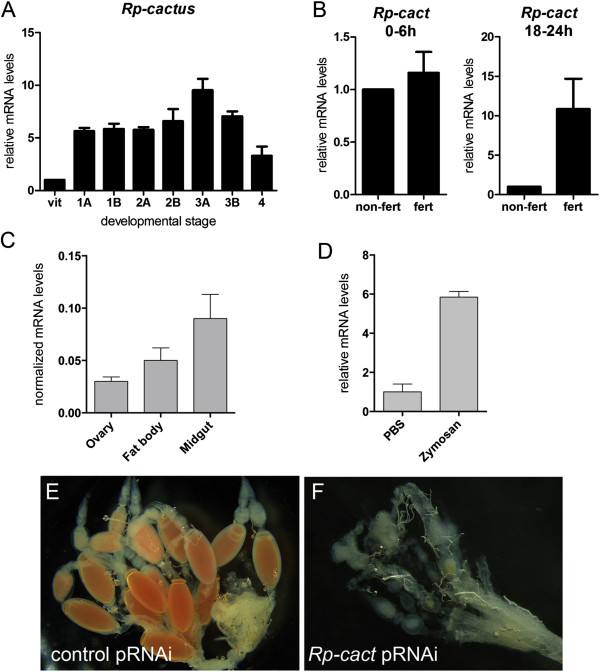
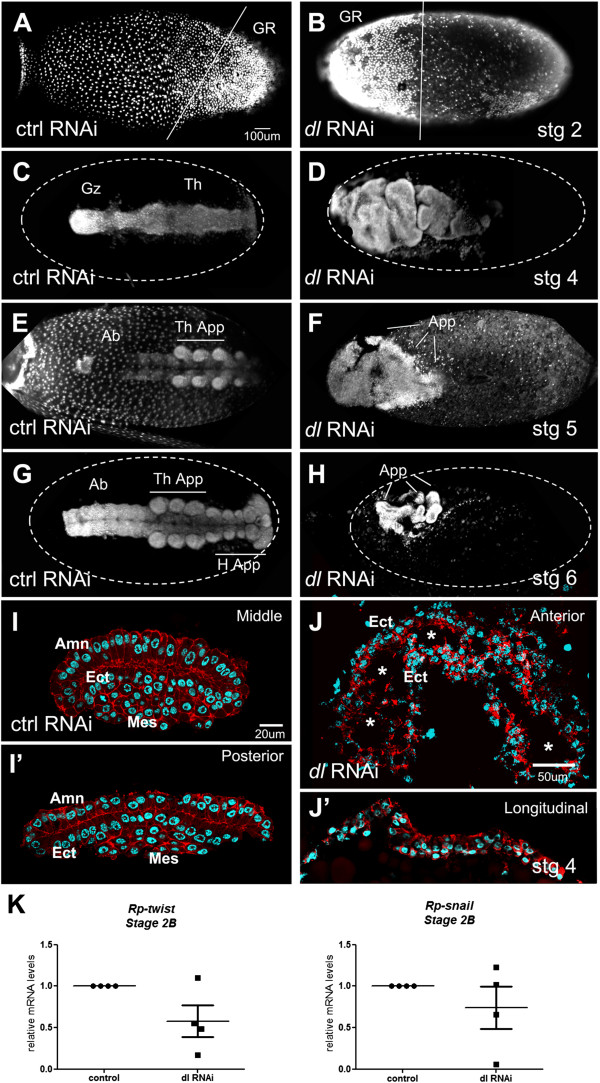
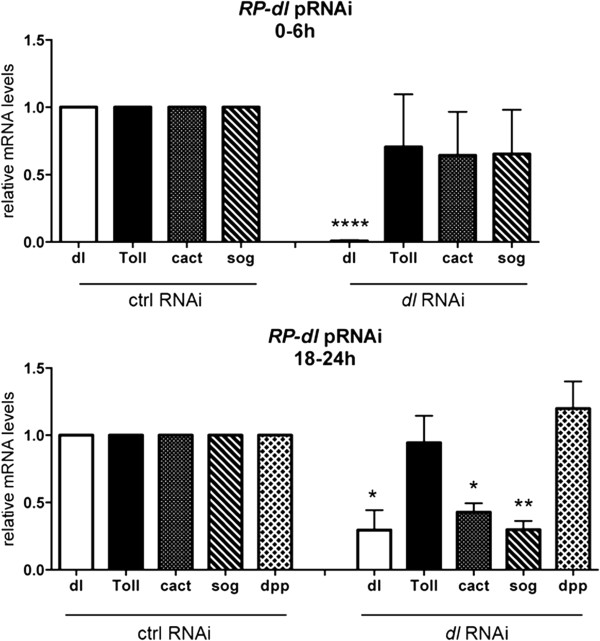
Results: We establish a staging system for early embryogenesis that allows us to correlate embryo morphology with gene expression profiles. Using this system, we investigate the role of Toll pathway genes during embryogenesis. Detailed analyses of gene expression throughout development, coupled with functional analyses using parental RNA interference, revealed that maternal Toll is required to establish germ layers along the dorso-ventral axis and for embryo placement along the anterior-posterior axis. Interestingly, knockdown of the Toll pathway effector Rp-dorsal appears to regulate the expression of the Bone Morphogenetic Protein antagonist Rp-short-gastrulation.
Conclusions: Our results indicate that Toll signals are the initiating event in dorso-ventral patterning during Rhodnius embryogenesis, and this is the first report of a conserved role for Toll in a hemipteran. Furthermore, as Rp-dorsal RNA interference generates anteriorly misplaced embryos, our results indicate a novel role for Toll signals in establishment of the anterior-posterior axis in Rhodnius.
Keywords: Dorso-ventral axis; Embryogenesis; Hemipteran; NFκB; Toll.
Figures
References
-
- Chen G, Handel K, Roth S. The maternal NF-kappaB/dorsal gradient of Tribolium castaneum: dynamics of early dorsoventral patterning in a short-germ beetle. Development. 2000;127:5145–5156. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
