

Genome Protection by the 9-1-1 Complex Subunit HUS1 Requires Clamp Formation, DNA Contacts, and ATR Signaling-independent Effector Functions
- PMID: 25911100
- PMCID: PMC4463431
- DOI: 10.1074/jbc.M114.630640
Genome Protection by the 9-1-1 Complex Subunit HUS1 Requires Clamp Formation, DNA Contacts, and ATR Signaling-independent Effector Functions
Abstract
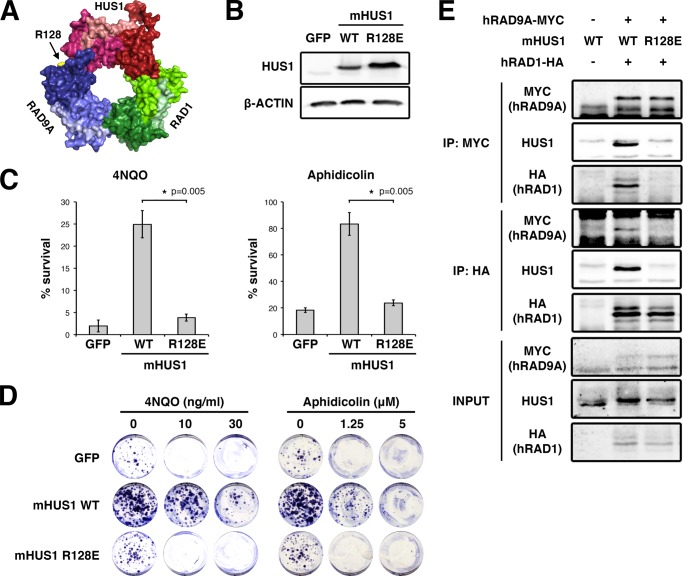
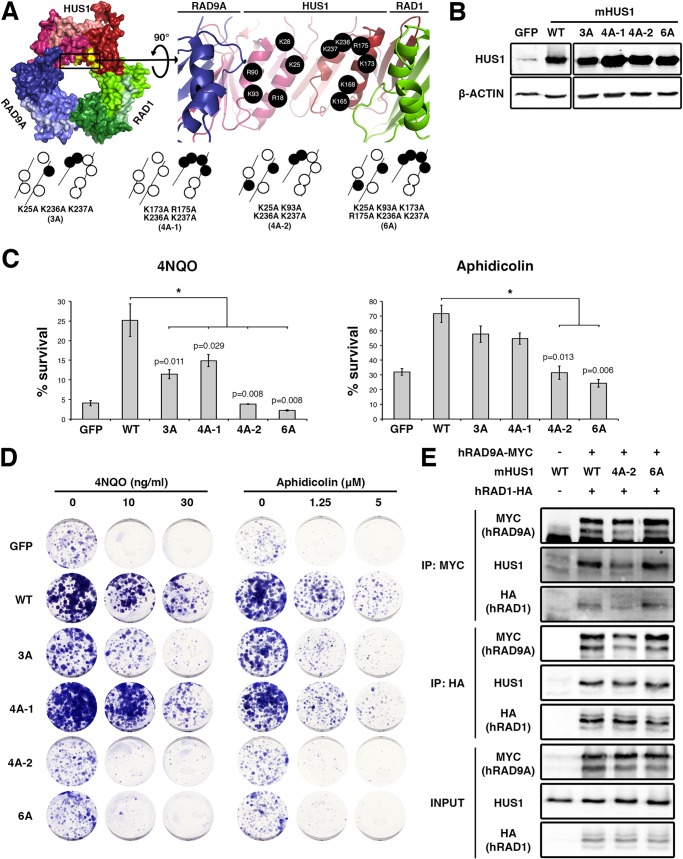
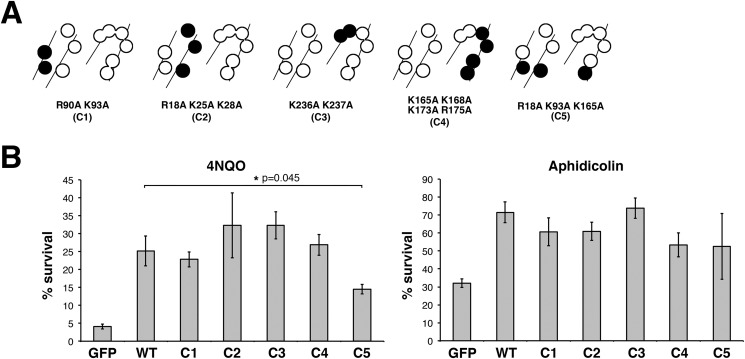
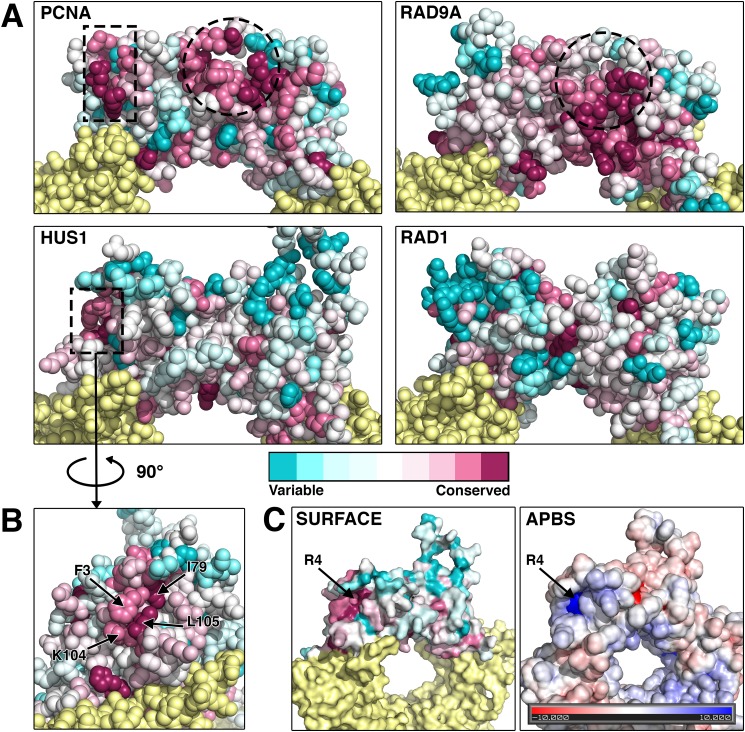
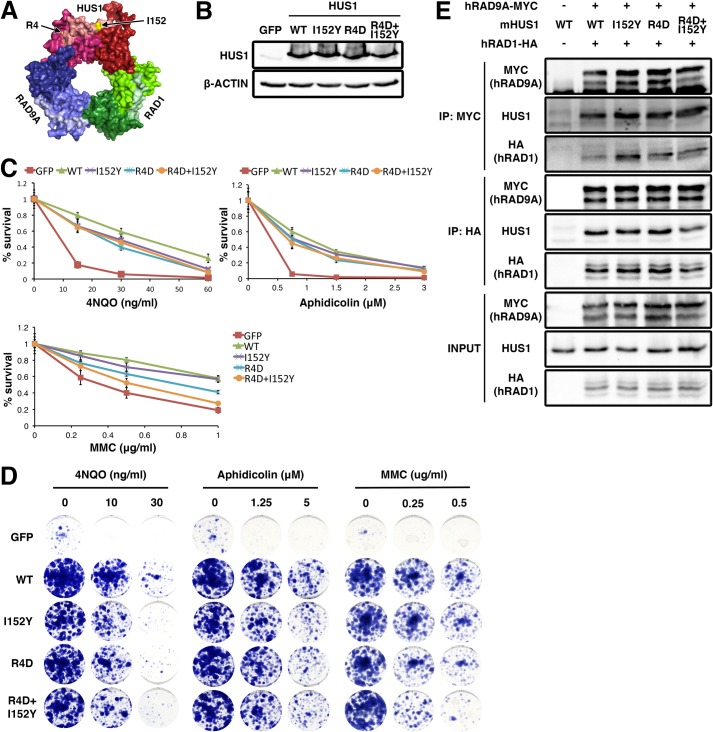
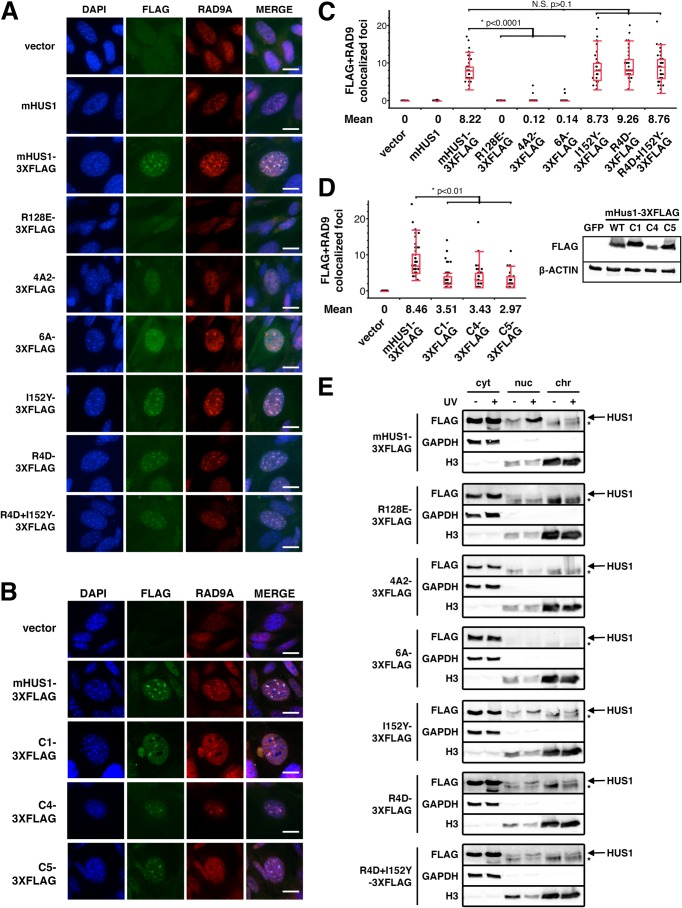
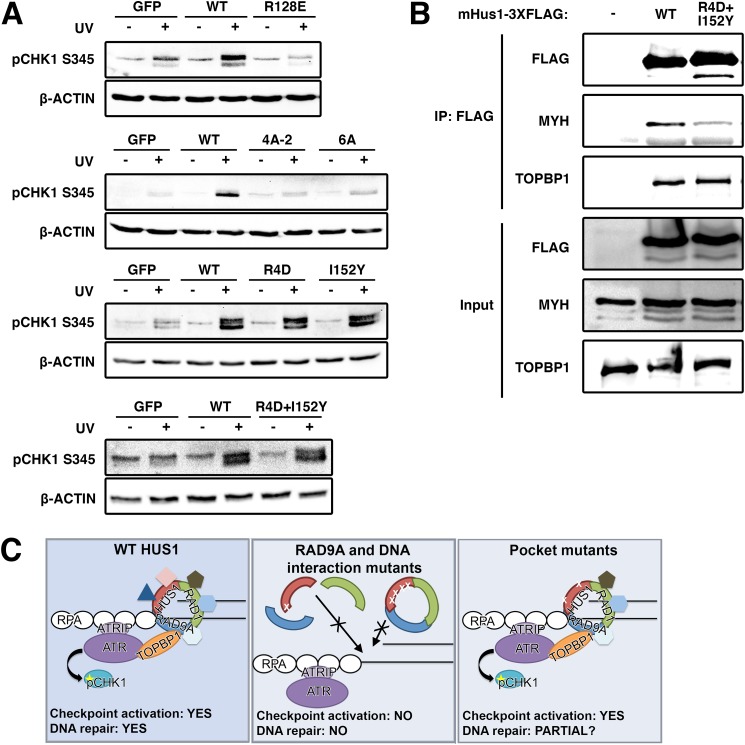
The RAD9A-HUS1-RAD1 (9-1-1) complex is a heterotrimeric clamp that promotes checkpoint signaling and repair at DNA damage sites. In this study, we elucidated HUS1 functional residues that drive clamp assembly, DNA interactions, and downstream effector functions. First, we mapped a HUS1-RAD9A interface residue that was critical for 9-1-1 assembly and DNA loading. Next, we identified multiple positively charged residues in the inner ring of HUS1 that were crucial for genotoxin-induced 9-1-1 chromatin localization and ATR signaling. Finally, we found two hydrophobic pockets on the HUS1 outer surface that were important for cell survival after DNA damage. Interestingly, these pockets were not required for 9-1-1 chromatin localization or ATR-mediated CHK1 activation but were necessary for interactions between HUS1 and its binding partner MYH, suggesting that they serve as interaction domains for the recruitment and coordination of downstream effectors at damage sites. Together, these results indicate that, once properly loaded onto damaged DNA, the 9-1-1 complex executes multiple, separable functions that promote genome maintenance.
Keywords: DNA damage, DNA damage response, checkpoint control, proliferating cell nuclear antigen (PCNA), genomic instability, HUS1, 9-1-1 complex.
© 2015 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Eichinger C. S., Jentsch S. (2011) 9-1-1: PCNA's specialized cousin. Trends Biochem. Sci. 36, 563–568 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
